植物糖生物学研究进展_尹恒

植物学报 Chinese Bulletin of Botany 2010, 45 (5): 521–529, https://www.360docs.net/doc/4c3467283.html, doi: 10.3969/j.issn.1674-3466.2010.05.001
——————————————————
收稿日期: 2010-01-18; 接受日期: 2010-03-23
基金项目: 863计划(No.2006AA10A213, No.2007AA091601)和中国科学院知识创新工程重要方向项目(No. KSCX2-YW-G-041) * 通讯作者。E-mail: zxm@https://www.360docs.net/doc/4c3467283.html,; dyguang@https://www.360docs.net/doc/4c3467283.html,
植物糖生物学研究进展
尹恒, 王文霞, 赵小明*, 杜昱光*
中国科学院大连化学物理研究所辽宁省碳水化合物重点实验室, 大连 116023
摘要 自1988年糖生物学概念提出以来, 国内外科学家在动物、微生物领域取得了大量的研究成果, 但植物糖生物学的研究进展较慢, 目前少见系统的专著或综述。该文围绕植物正常生长时糖信号、逆境时糖信号、糖蛋白及其糖链、重要糖基转移酶及植物凝集素等植物糖生物学的主要问题, 全面阐述植物糖生物学的各个研究分支, 并介绍各领域的最新研究进展。提出了植物糖生物学的概念, 并将其定义为研究植物与糖类互作机制及植物体内糖(糖链与糖分子)结构及生物学功能的科学。
关键词 糖蛋白, 糖基转移酶, 凝集素, 植物糖生物学, 糖信号
尹恒, 王文霞, 赵小明, 杜昱光 (2010). 植物糖生物学研究进展. 植物学报 45, 521–529.
糖类是生物体的重要组成成分, 在自然界中分布广泛, 含量丰富。但直到20世纪上半叶, 糖类仍被视为是缺乏生物特异性的一类惰性化合物, 只是作为代谢能量来源或充当结构保护材料(如植物细胞壁和昆虫的外壳), 在生物体内功能较少。由于糖类物质结构复杂、糖链分析技术缺乏, 科学家们对其研究关注不多, 使得糖类的研究远远落后于另2种生物大分子 ——核酸和蛋白质。
20世纪70年代以来, 随着糖链解析技术水平的提高以及分子生物学的发展, 尤其是人、拟南芥(Arabidopsis thaliana )等模式生物基因组测序的完成, 围绕糖类物质的研究工作日渐增多。越来越多的证据表明, 糖类物质全面参与了生物的生殖发育、生长、应激等过程, 是很多生理和病理过程中分子识别的决定因素。最初, 这些围绕糖的研究工作被认为是糖化学的一个分支, 但很快其中大量的生物学工作远远超出了糖化学的范畴, 因此科学家们提出了糖生物化学的概念, 而随着研究内容的进一步深入, 糖生物化学也不能完全涵盖糖在生物领域的最新研究进展。1988年, 生化领域的著名杂志《生物化学年评》发表了英国牛津大学Rademacher 等人题为“糖生物学(Glycobiology)”的一篇综述文章(Rademacher et al., 1988), 标志着糖生物学这一学科的正式诞生。此后,
围绕着糖链结构及糖的生物学功能, 科学家们在糖链与疾病的关系、天然产物中糖的分离提纯以及功能糖的制备与应用等方面进行了大量的工作, 取得了一定进展。2001年, Science 杂志汇编了Hurtley 等人的7篇综述和6篇简介, 以《灰姑娘的马车来了》为题编辑了一期“糖和糖生物学”专辑, 对糖生物学最新的研究成果及前景进行了综述和展望, 从而将糖生物学的研究推向了一个新的高度(Hurtley et al., 2001)。2006年, Nature 杂志也推出了糖化学与糖生物学的专辑, 全面介绍了糖生物学领域的研究进展。我国糖生物学的开展与国际接轨较快, 1995年金城等人将糖生物学概念引入中国(金城和张树政, 1995), 此后, 我国科学家在糖生物合成和糖链功能解析等领域取得了一定进展。
广义糖生物学的含义是: 研究自然界中广泛分布的糖(糖链或聚糖)的结构、生物合成和生物学意义。但有关糖类结构和生物合成的研究也是已有学科糖化学和糖生物化学的主要研究内容之一, 所以糖生物学研究和讨论的对象更多地聚焦在一些重要的功能糖、生物体内糖缀合物的生物学功能上。实际上, 糖生物学的研究焦点是糖类和其它分子的关系, 有一种观点认为, 蛋白质和糖类的相互作用是糖生物学的基础(王克夷, 2009)。目前糖生物学的工作多围绕动物、
·特邀综述·
522 植物学报 45(5) 2010
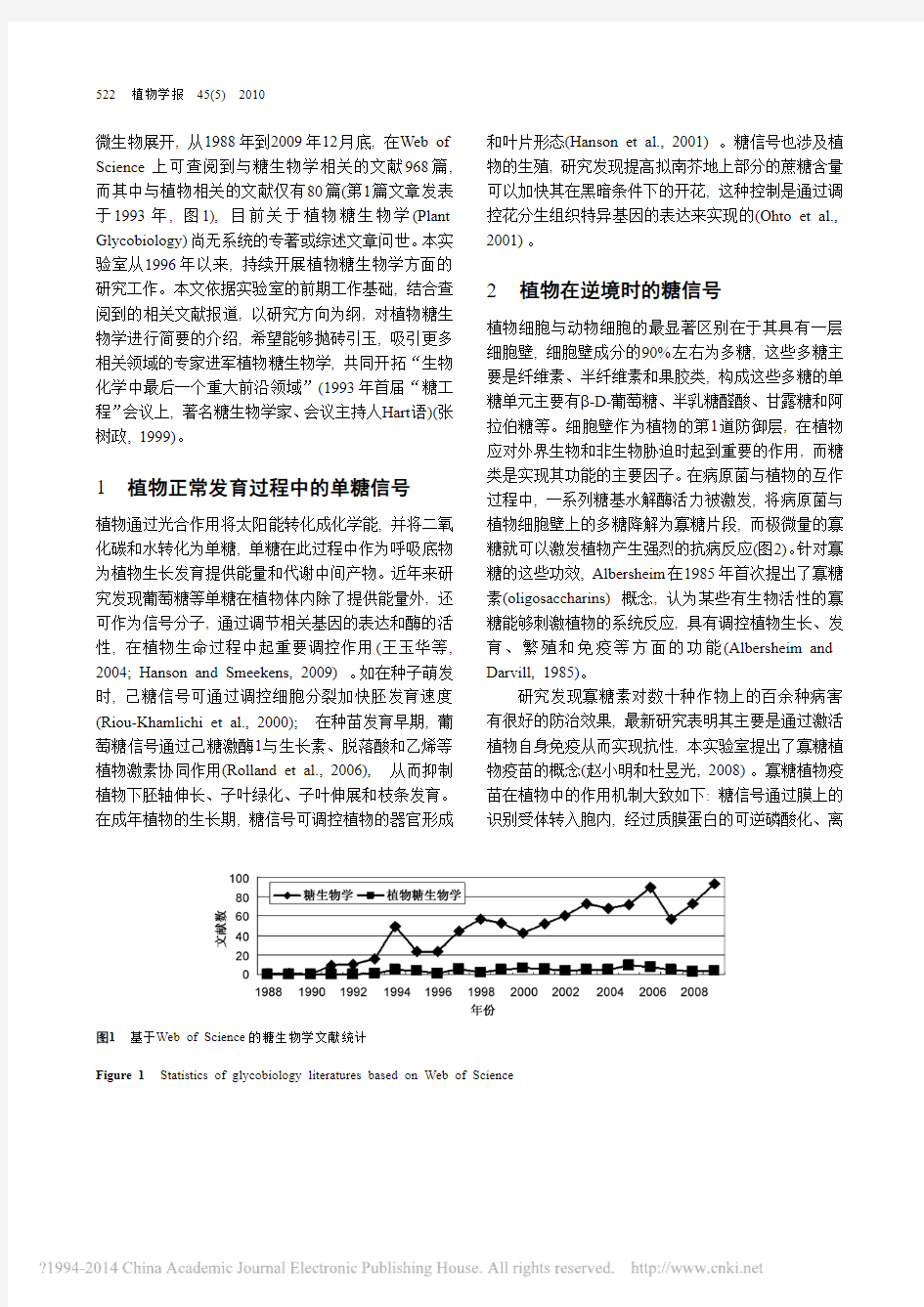
微生物展开, 从1988年到2009年12月底, 在Web of Science上可查阅到与糖生物学相关的文献968篇, 而其中与植物相关的文献仅有80篇(第1篇文章发表于1993年, 图1), 目前关于植物糖生物学(Plant Glycobiology)尚无系统的专著或综述文章问世。本实验室从1996年以来, 持续开展植物糖生物学方面的研究工作。本文依据实验室的前期工作基础, 结合查阅到的相关文献报道, 以研究方向为纲, 对植物糖生物学进行简要的介绍, 希望能够抛砖引玉, 吸引更多相关领域的专家进军植物糖生物学, 共同开拓“生物化学中最后一个重大前沿领域”(1993年首届“糖工程”会议上, 著名糖生物学家、会议主持人Hart语)(张树政, 1999)。
1植物正常发育过程中的单糖信号
植物通过光合作用将太阳能转化成化学能, 并将二氧化碳和水转化为单糖, 单糖在此过程中作为呼吸底物为植物生长发育提供能量和代谢中间产物。近年来研究发现葡萄糖等单糖在植物体内除了提供能量外, 还可作为信号分子, 通过调节相关基因的表达和酶的活性, 在植物生命过程中起重要调控作用(王玉华等, 2004; Hanson and Smeekens, 2009)。如在种子萌发时, 己糖信号可通过调控细胞分裂加快胚发育速度(Riou-Khamlichi et al., 2000); 在种苗发育早期, 葡萄糖信号通过己糖激酶1与生长素、脱落酸和乙烯等植物激素协同作用(Rolland et al., 2006), 从而抑制植物下胚轴伸长、子叶绿化、子叶伸展和枝条发育。在成年植物的生长期, 糖信号可调控植物的器官形成和叶片形态(Hanson et al., 2001)。糖信号也涉及植物的生殖, 研究发现提高拟南芥地上部分的蔗糖含量可以加快其在黑暗条件下的开花, 这种控制是通过调控花分生组织特异基因的表达来实现的(Ohto et al., 2001)。
2植物在逆境时的糖信号
植物细胞与动物细胞的最显著区别在于其具有一层细胞壁, 细胞壁成分的90%左右为多糖, 这些多糖主要是纤维素、半纤维素和果胶类, 构成这些多糖的单糖单元主要有β-D-葡萄糖、半乳糖醛酸、甘露糖和阿拉伯糖等。细胞壁作为植物的第1道防御层, 在植物应对外界生物和非生物胁迫时起到重要的作用, 而糖类是实现其功能的主要因子。在病原菌与植物的互作过程中, 一系列糖基水解酶活力被激发, 将病原菌与植物细胞壁上的多糖降解为寡糖片段, 而极微量的寡糖就可以激发植物产生强烈的抗病反应(图2)。针对寡糖的这些功效, Albersheim在1985年首次提出了寡糖素(oligosaccharins)概念, 认为某些有生物活性的寡糖能够刺激植物的系统反应, 具有调控植物生长、发育、繁殖和免疫等方面的功能(Albersheim and Darvill, 1985)。
研究发现寡糖素对数十种作物上的百余种病害有很好的防治效果, 最新研究表明其主要是通过激活植物自身免疫从而实现抗性, 本实验室提出了寡糖植物疫苗的概念(赵小明和杜昱光, 2008)。寡糖植物疫苗在植物中的作用机制大致如下: 糖信号通过膜上的识别受体转入胞内, 经过质膜蛋白的可逆磷酸化、离
图1基于Web of Science的糖生物学文献统计
Figure 1 Statistics of glycobiology literatures based on Web of Science
尹恒等: 植物糖生物学研究进展
523
图2 植物与病原菌互作中产生的寡糖素信号(http://www.gak. co.jp/FCCA/glycoword/SA-A01/SA-A01_E.html)
Figure 2 Oligosaccharide signaling in plant-pathogen inter- actions (http://www.gak.co.jp/FCCA/glycoword/SA-A01/SA- A01_E.html)
子通道变化、质膜去极化、活性氧和一氧化氮爆发、植物激素(水杨酸、茉莉酸、乙烯等)产生、丝裂原活化蛋白激酶信号通路激活等一系列信号转导和放大过程, 调控相关防卫基因的表达, 促进抗性次生代谢产物积累, 诱导抗性反应产生, 从而最终有效地抵抗病原物侵染(Garcia-Brugger et al., 2006)。
在此基础上, 科学家们还发现, 许多糖类物质具有促进植物生长、提高产品品质和抵御自然灾害(如冷冻、干旱)的功效。如来源于海带中的海藻酸及其寡糖在白菜、烟草等多种作物上有良好的促进生长作用。最新的研究发现, 来源于海洋虾蟹壳的壳寡糖用于苹果、梨、芒果和葡萄等水果时, 具有良好的抗冻、保花护果及改善品质的效果。这些寡糖类物质因为其绿色天然、安全高效和环境友好等特点, 被认为是一种新型的农用制剂, 在我国的绿色农业生产中将发挥重要作用。
在根瘤菌与豆科植物的互作过程中存在一种特殊的糖类信号——结瘤因子(nod factor)。结瘤因子是根瘤菌在宿主植物根系分泌的类黄酮的作用下合成并分泌的脂几丁寡糖(lipochito-oligosaccharide)信号分子, 一般由一个3–5个N-乙酰氨基葡萄糖组成的糖骨架及其非还原端C 2上的一条脂肪酸链构成, 在根
瘤菌与植物的共生固氮过程中起重要作用。结瘤因子被植物中的特定受体“结瘤因子结合蛋白”识别(Geurts and Bisseling, 2002), 再激活Ca 2+介导的信号转导途径, 最终诱导宿主植物根毛变形、皮层细胞分裂及结瘤素基因表达等一系列反应从而实现功能(Geurts et al., 2005)。结瘤因子上几丁寡糖骨架的长度、脂肪酸的结构以及还原端和非还原端的化学修饰都会影响结瘤因子的活性(D'Haeze and Holsters, 2002), 这也说明了糖信号的复杂性和特异性。
3 植物糖蛋白
生物体内有1/3以上的蛋白质是糖蛋白, 糖蛋白功能是目前动物糖生物学研究中最活跃的领域。糖蛋白被定义为由糖链与蛋白质分子共价相连构成的一类复合物, 这些糖链被称为寡糖链(oligosaccharide chain)或聚糖(glycan), 由少则1个、多则数百个糖基连成。构成植物糖蛋白的主要糖基有半乳糖、N-乙酰氨基葡萄糖、甘露糖、阿拉伯糖和木糖等。
植物糖蛋白广泛存在于细胞壁、细胞膜和细胞质中, 包括许多酶、凝集素、结构蛋白质、贮藏蛋白质以及受体, 在植物的种子萌发、幼苗生长、生殖和应激等生命过程中均起重要的作用。目前已鉴定得到多个重要的植物糖蛋白家族, 如富含羟脯氨酸糖蛋白(与植物诱抗相关)和阿拉伯半乳糖蛋白(在被子植物受精过程中起作用)等。尤其是细胞表面糖蛋白, 往往是最先接受外界信号的受体, 负责细胞与外界的信息交换, 所以科学家们更重视细胞表面糖蛋白的功能研究。2006年, 日本的Shibuya 实验室鉴定得到了几丁寡糖在水稻(Oryza sativa )质膜上的受体蛋白CEBiP, 结构分析表明此受体是一个具有11个糖基化位点的糖蛋白, 电泳分析显示CEBiP 由35 kDa 的蛋白质和40 kDa 的糖链组成(Kaku et al., 2006)。而其它一些激发子鞭毛蛋白等的受体也是糖蛋白(Chinchilla et al., 2006)。
糖蛋白中蛋白质是生理功能的主要担负者, 而糖链则对蛋白质的功能起辅助修饰作用, 糖链对蛋白质功能的主要影响实际上是通过影响蛋白质的整体构象实现的。糖蛋白中糖链的作用有2大类: 分子内的作用(如蛋白质的正确折叠、细胞内定位等)和分子间的作用(如细胞粘附和结合病原体等)。研究发现糖链
524 植物学报 45(5) 2010
结构组成的微小差异也会导致其功能的显著差异, 糖链功能与其单糖组成和数目、排列顺序和构型、糖链的长短及连接位置均有紧密联系。但糖链的结构复杂(如由3个不同核苷酸残基或3个不同氨基酸残基组成的直链分子只有6种可能的序列, 而由3个己糖基组成的聚糖, 其可能的序列有1 056–27 648种), 研究难度很大; 而且同样结构的糖链连接在不同蛋白上其功能不同; 即使是在同一个蛋白上, 其在生物体的不同部位和不同的发育阶段也有不同的功能。因而对于植物糖蛋白上各类糖链的特定功能, 目前还没有发现较好的规律, 对糖链的生物学作用仍只能逐个单独研究。另外, 植物糖蛋白上糖链的结构分析也是植物糖生物学的一个重要研究分支, 目前通常使用高效液相色谱(HPLC)和质谱串联技术进行糖链结构解析, 首先通过对糖链进行柱前衍生化带上紫外或荧光基团后经由HPLC分离, 得到纯化的糖链样品后再利用电喷雾质谱或飞行质谱进行结构解析(王仲孚等, 2006)。但由于植物体系比较复杂, 对植物中糖蛋白糖链的结构解析与功能研究报道很少(Seveno et al., 2008)。日本的Kimura实验室在此方面进行了大量的工作, 1997年, 他们结合荧光标记、高效排阻液相色谱和离子喷雾串联质谱等多种手段鉴定了大豆(Glycine max)种子中8个贮藏蛋白质上的糖链结构, 此后, 科学家们仅对烟草(Nicotiana tabacum)、花生(Arachis hypogaea)和番茄(Lycopersicon esculen-tum)等植物中的少数糖蛋白成功进行了糖链结构解析(Kimura et al., 2008)。值得一提的是, 1995年, 我国上海有机化学研究所的田庚元研究员也曾利用全水解和部分酸水解、甲基化分析、CC、CC/MS、1H 和12C-NMR等方法解析了枸杞子(Lycium barbarum)中糖蛋白的糖链结构(田庚元等, 1995)。
4糖基转移酶
糖蛋白是通过蛋白质的糖基化组装实现的, 而糖基化过程则通过多种糖基转移酶完成——在肽链合成的同时或合成后, 在糖基转移酶的催化下, 糖链被连接到肽链的特定糖基化位点上。糖基转移酶具有高度的底物专一性, 即同时对糖基的供体和受体具有专一性。对糖基转移酶进行研究, 是糖基化研究的第1步, 有助于了解蛋白质上的寡糖链, 为揭示糖链及糖蛋白功能奠定基础。因此, 对糖基转移酶的研究成为糖生物学的一个重要分支, 而由于其研究手段相对较多, 也是目前糖生物学中开展最好的研究领域。下文简要介绍几类重要的糖基转移酶。
4.1 N-乙酰氨基葡萄糖转移酶(N-acetylglucosa- minyl-transferase, GnT)
糖蛋白中糖链通过还原端的N-乙酰氨基葡萄糖(GlcNAc)以β-1,4糖苷键与蛋白质肽链上Asn- XXX-Ser/Thr序列(XXX为除脯氨酸以外的氨基酸)中Asn残基上的氨基(-NH2)相连, 被称为N-糖链。真核细胞中N-糖链的合成途径高度保守, 其第1步合成由GnT完成。1999年, Strasser等依据动物GnT保守区序列设计简并引物, 从烟草文库中分离到编码GnT的基因GnTI (Strasser et al., 1999), 这也是植物中第1个被鉴定的GnT基因。随后利用同样的方法从拟南芥、马铃薯(Solanum tuberosum)中分离和鉴定出一系列GnT基因, 这些基因与动物GnT基因均有较高的序列相似性。后续研究发现GnTI定位于植物的内质网和高尔基体(Saint-Jore-Dupas et al., 2006), 而减弱了GnTI活力的植株并不对其下游的其它糖基转移酶活力构成影响(Strasser et al., 2004), 说明植物体内具有GnTI的功能类似物。
4.2 多肽N-乙酰氨基半乳糖转移酶(polypeptide- N-acetylgalactosaminyltransferase, ppGalNAc) O-连接糖链有多种形式, 在动物中研究最深入的是O-乙酰氨基半乳糖(O-GalNAc)连接糖链, 该糖链通过还原端的N-乙酰氨基半乳糖(GalNAc)以β-1,4糖苷键与蛋白质肽链上Ser或Thr的氧原子连接。ppGalNAc催化O-GalNAc连接糖链合成的第1步, 将UDP-GalNAc上的GalNAc基团转移至多肽链上特定序列中的Ser或Thr的羟基上, 从而合成GalNAc- O-Ser/Thr糖蛋白片段。但植物中与O-GalNAc连接糖链相关的研究十分罕见, 仅有2篇报道: 1999年Kishimoto等人报道水稻的谷蛋白中含有O-GalNAc 连接糖链(Kishimoto et al., 1999); 最近Kilcoyne等(2009)发现在醇溶的水稻蛋白中也存在O-GalNAc连接的糖蛋白, 但尚未鉴定出此蛋白质。目前研究者尚未得到植物中的任何ppGalNAc基因, 这也将是今后开展植物糖生物学研究的一个较好切入点。
尹恒等:植物糖生物学研究进展 525
4.3 O-乙酰氨基葡萄糖糖基转移酶(O-GlcNAc transferase, OGT)
与前述2种糖基化多发生在内质网和高尔基体不同, 近年来在细胞核和细胞质中发现存在另一种O连接N-乙酰氨基葡萄糖糖基化(O-GlcNAc)过程。O-GlcNAc糖基化修饰在细胞内分布广泛, 指通过OGT将单个N-GlcNAc添加到蛋白质的Ser或Thr残基上。这种作用与蛋白磷酸化作用类似, O-GlcNAc修饰很可能通过改变蛋白的细微结构或形成空间位阻从而抑制该肽链临近位置的磷酸化, 从而共同参与转录调控、信号转导等生命活动。这种被称作“阴阳调控”的关系在全细胞水平和特定蛋白的特定位点上都已得到验证(Hart et al., 1995)。
美国明尼苏达大学的Olszewski研究组长期从事植物中OGT酶的研究工作。1996年他们首先从拟南芥中克隆得到了植物中的第1个OGT基因——SPINDLY(SPY)(Jacobsen et al., 1996)。SPY与动物的OGT具有较高的同源性, 含有34肽重复序列, 由作为信号域的TRP结构域和催化功能域2部分组成。SPY蛋白在植物细胞质与细胞核部位都有出现, 但大部分出现在后者, 而它不具有明显的核定位信号, 因此与其它具有核定位信号的蛋白相互作用可能是SPY定位于细胞核的原因。在此之后, 大麦(Hordeum vulgare)、矮牵牛(Petunia hyhrida)、番茄和水稻中的OGT基因也被陆续发现, 它们与拟南芥SPY均具有较高同源性, 说明O-GlcNAc修饰在植物中极度保守。最初研究认为, SPY仅在赤霉素信号途径中起负调控因子的作用, 但后续研究发现, SPY还可影响植物中的脱落酸、油菜素内酯等信号通路(Shimada et al., 2006), 并调控拟南芥中的活性氧爆发(Cao et al., 2006)。表明植物OGT不仅在植物发育过程中起作用, 它还涉及植物抗性反应。而SPY的RNAi植株呈现致死性(Hartweck et al., 2002), 进一步说明了O-GlcNAc糖基化在植物生理活动中的重要性。
4.4 植物小分子糖苷糖基转移酶
除了将糖转移到肽链上, 在植物体中, 糖基转移酶还可将糖基转运到激素和脂类等小分子上。这类反应的功能包括改变代谢物的水溶性、稳定性、运输特性、结合特性、调控亚细胞定位以及消解有害代谢物或环境化合物毒性等(Wang, 2009)。目前已经从植物中鉴定出数十种具有此种功能的糖基转移酶。
植物中小分子糖苷的糖基化影响到植物的全部生命过程, 在植物激素调控、次生代谢、信号转导和防卫反应等领域都起到重要的作用。尤其是其对植物激素的修饰, 起到了控制激素平衡的重要作用。在植物体内除乙烯外, 其余激素的糖基化衍生物均已被发现, 这些糖苷物被认为在植物体内起到了激素库的作用(Gachon et al., 2005)。我国山东大学的侯丙凯教授在此领域取得了丰硕的成果, 2004年他们发现拟南芥中的细胞分裂素可被糖基转移酶在不同位点修饰形成无活性的N-糖苷细胞分裂素和O-糖苷细胞分裂素; 前者是细胞分裂素的永久失活形式, 后者是细胞分裂素的贮藏形式, 在一定条件下可通过去糖基化而恢复细胞分裂素活性(Hou et al., 2004)。对植物中小分子化合物的糖基化及相关糖基转移酶的研究将有助于了解植物体内激素的平衡模式从而拓宽我们对植物生命活动的了解。
5凝集素
凝集素是一类具有高度特异性糖结合活性的糖蛋白, 动物和微生物的凝集素被认为是糖信号的“解密器”, 在动物先天免疫、细胞周期调节等过程中起作用。植物凝集素存在于许多植物的种子和营养器官中。1888年, 第1个植物凝集素在蓖麻籽中被发现, 之后多种植物凝集素不断被发现。到目前为止, 已经发现了1 000多种不同的植物凝集素。植物凝集素被定义为至少具有1个可与糖分子特异可逆结合的非催化结构域的植物蛋白, 但一般凝集素上都有多个与糖链结合的位点, 植物凝集素具有较高的同源性, 可通过保守区域比对筛选找到新的凝集素(Shridhar et al., 2009)。凝集素在结合糖的过程中, 表面局部结构的构象会有所变化, 这种变化有利于其识别并结合外界糖信号。根据凝集素专一识别糖类的不同, 可以大致分为6个组别: 岩藻糖组、半乳糖/N-乙酰半乳糖胺组、N-乙酰葡萄糖胺组、甘露糖组、唾液酸组和复合糖组。与其它糖蛋白类似, 凝集素的功能是通过其分子中肽链的活性部位, 即专一结合糖的区域(CRD)实现的; 而与凝集素上的糖链关系不大, 糖链主要起到维持凝集素构型的作用。
526 植物学报 45(5) 2010
由于植物凝集素广泛存在于植物的不同组织器官及各个发育阶段, 同时有特定的糖结合专一性, 因此, 凝集素在植物中的生理功能是多种多样的。较一致的意见是凝集素的糖结合活性及其专一性决定了它们的功能。因为植物凝集素对许多外源多糖有明显的结合作用, 最初人们多研究其外源作用: 毒性凝集素可以起到防御食草动物和昆虫侵蚀的作用, 被侵蚀后, 植物凝集素就会从植物的受害细胞中释放出来, 进入危害者的消化系统, 与特定的糖结合引发毒性, 抑制动物及昆虫的生长繁殖甚至致死(Vandenborre et al., 2009)。同样, 植物凝集素可通过与病原菌的细胞壁结合抑制病原菌的生长(如几丁凝集素可与真菌细胞壁的几丁质结合), 从而起到抗病的作用; 此类凝集素基因可被转入相应植物中, 构建抗病虫害转基因品系(Ye et al., 2009), 为高效农业奠定基础。此外, 植物凝集素还参与植物与外界真菌的共生作用, 如豆科植物根部的凝集素可参与对Rhizobium的识别和结合, 在固氮过程中起重要作用(Laus et al., 2006)。
后来人们认识到植物凝集素不仅对外源信号起作用, 在植物体内也有丰富的生理功能。凝集素可以与植物的贮存蛋白结合, 调控其降解过程; 凝集素还可以与糖基水解酶和磷酸酶结合并影响它们的活性(Rudiger and Gabius, 2001), 这个现象说明凝集素可以调节植物体内的代谢及信号转导, 其作用领域远比人们预想的要更为广泛。最新研究发现在植物中还具有一类含有凝集素结构域(lectin domain)的激酶, 这类蛋白同时具有糖结合和磷酸化的功能, 在植物的生长和应激过程中具有重要作用(Bouwmeester and Govers, 2009)。更为有趣的是, 凝集素还参与了对植物蛋白糖基化的调节。中国科学院植物研究所种康研究组发现小麦(Triticum aestivum)春化过程中其总蛋白的O-GlcNAc糖基化水平显著提高, 而凝集素VER2通过磷酸化对此O-GlcNAc糖基化过程进行调控(Xing et al., 2009)。此工作涉及植物糖生物学的2个方面(蛋白糖基化与凝集素), 权威的生物学文献评价网站F1000Biology的述评认为此文对揭示植物的春化作用机制具有重要意义(http://f1000biology. com/article/id/1163821/evaluation)。
除上述作用外, 因为凝集素具有与糖类专一性结合的特性, 可作为糖类物质的探针分子广泛应用于糖生物学研究。凝集素可简单用于糖的鉴定; 也可被用于亲和色谱及毛细管电泳中进行糖的分离; 还可利用凝集素与糖链的亲和力研制生物传感器(Yang et al., 2006)。最近, 有研究者将凝集素固定化制作成微阵列芯片, 可高通量地检测整个细胞表面所表达的糖基类型(Wu et al., 2009)。
6 现状与展望
虽然糖类物质结构复杂, 研究难度较大, 但自从1988年糖生物学概念提出以来, 糖生物学的研究已在世界各国引起了高度的关注并取得了令人惊喜的进展。日本的“糖工程前沿计划”、美国的“功能糖组学”等大规模研究计划纷纷上马; 英国牛津大学糖生物学研究所等优秀科研机构脱颖而出; 创办了Glycobiology 等优秀的专业期刊; 还形成了3个每年固定召开的学术会议——戈登糖生物学年会、糖生物学年会和糖复合物研讨会(第22届会议将于2011年在我国大连召开)。
目前糖生物学大部分的研究成果是在动物和微生物上获得的, 在植物糖生物学领域研究工作还不够深入, 有很多问题尚未解决。但随着糖生物学概念的深入和推广, 随着科研人员的重视, 我们相信植物糖生物学的研究将迎来高速发展时期。依据上文介绍的研究进展, 我们将植物糖生物学定义为研究植物与糖类互作机制及植物体内糖(糖链与糖分子)结构与生物学功能的科学。从重要性与可行性出发, 笔者认为现阶段植物糖生物学的研究重点有以下3方面: 糖信号网络中关键节点的寻找, 植物糖蛋白上糖链的结构及功能解析和植物糖基转移酶的鉴定及功能研究。由于糖类物质在自然界的广泛存在性及糖基化在植物体内的普遍性, 植物糖生物学是全面揭示植物生命活动的重要研究领域, 希望更多的科研工作者能投身于此, 推动我国植物糖生物学研究工作, 力争在国际上抢占学术领先地位。
致谢作者尹恒的部分工作受到植物病虫害生物学国家重点实验室开放基金(No.SKL2010OP16)的资助; 中国科学院上海生物化学研究所王克夷研究员审阅本文, 并提出了许多中肯的建议, 在此一并表示感谢。
参考文献
金城, 张树政 (1995). 糖生物学与糖工程的兴起与前景. 生物
尹恒等:植物糖生物学研究进展 527
工程进展3,12–17.
田庚元, 王晨, 冯宇澄 (1995). 枸杞子糖蛋白的分离纯化、物化性质及糖肽键特征. 生物化学与生物物理学报2, 201–206.
王克夷 (2009). 糖生物学和糖组学. 生命的化学(3), 4–10.
王玉华, 杨清, 陈敏 (2004). 植物糖感知和糖信号传导. 植物学通报21, 273–279.
王仲孚, 贺建宇, 尉亚辉, 黄琳娟(2006). 用于寡糖链分析的HPLC柱前衍生化方法研究进展. 有机化学5, 592–598.
张树政 (1999). 糖生物学: 生命科学中的新前沿. 生命的化学
(3),3–7.
赵小明, 杜昱光(2008). 寡糖激发植物免疫及寡糖植物疫苗的研究进展. 见: 邱德文主编. 植物免疫与植物疫苗-研究与实践. 北京: 科学出版社. pp. 48–66.
Albersheim P, Darvill AG (1985). Oligosaccharins: novel molecules that can regulate growth, development, reproduction, and defense against disease in plants. Sci Am253,58–64.
Bouwmeester K, Govers F (2009). Arabidopsis L-type lectin receptor kinases: phylogeny, classification, and expression profiles. J Exp Bot60,4383–4396.
Cao SQ, Ye M, Huang Q, Zhang RX (2006). A role for SPINDLY gene in the regulation of oxidative stress response in Arabidopsis. Russ J Plant Physiol 53, 541–547.
Chinchilla D, Bauer Z, Regenass M, Boller T, Felix G (2006). The Arabidopsis receptor kinase FLS2 binds flg22 and determines the specificity of flagellin perception. Plant Cell18,465–476.
D'Haeze W, Holsters M (2002). Nod factor structures, responses, and perception during initiation of nodule development. Glycobiology12,R79–R105.
Gachon CMM, Langlois-Meurinne M, Saindrenan P (2005). Plant secondary metabolism glycosyltransferases: the emerging functional analysis. Trends Plant Sci10, 542–549.
Garcia-Brugger A, Lamotte O, Vandelle E, Bourque S, Lecourieux D, Poinssot B, Wendehenne D, Pugin A (2006). Early signaling events induced by elicitors of plant defenses. Mol Plant Microbe Interact 19,711–724.
Geurts R, Bisseling T (2002). Rhizobium nod factor perception and signaling. Plant Cell14,S239–S249. Geurts R, Fedorova E, Bisseling T (2005). Nod factor signaling genes and their function in the early stages of rhizobium infection. Curr Opin Plant Biol8,346–352. Hanson J, Johannesson H, Engstrom P (2001).
Sugar-dependent alterations in cotyledon and leaf development in transgenic plants expressing the HDZhdip gene ATHB13. Plant Mol Biol45,247–262.
Hanson J, Smeekens S (2009). Sugar perception and signaling—an update. Curr Opin Plant Biol12,562–567. Hart GW, Greis KD, Dong LYD, Blomberg MA, Chou TY, Jiang MS, Roquemore EP, Snow DM, Kreppel LK, Cole RN, Comer FI, Arnold CS, Hayes BK (1995). O-linked N-acetylglucosamine: The ''yin-yang'' of ser/thr phospho- rylation? Nuclear and cytoplasmic glycosylation.
Glycoimmunology376,115–123.
Hartweck LM, Scott CL, Olszewski NE (2002). Two O-linked N-acetylglucosamine transferase genes of Arabidopsis thaliana L. Heynh. have overlapping functions necessary for gamete and seed development.
Genetics 161, 1279–1291.
Hou BK, Lim EK, Higgins GS, Bowles DJ (2004).
N-glucosylation of cytokinins by glycosyltransferases of Arabidopsis thaliana. J Biol Chem279,47822–47832. Hurtley S, Service R, Szuromi P (2001). Cinderella's coach is ready. Science291,2337–2337.
Jacobsen SE, Binkowski KA, Olszewski NE (1996).
SPINDLY, a tetratricopeptide repeat protein involved in gibberellin signal transduction in Arabidopsis. Proc Natl Acad Sci USA93,9292–9296.
Kaku H, Nishizawa Y, Ishii-Minami N, Akimoto-Tomiyama C, Dohmae N, Takio K, Minami E, Shibuya N (2006).
Plant cells recognize chitin fragments for defense signaling through a plasma membrane receptor. Proc Natl Acad Sci USA103,11086–11091.
Kilcoyne M, Shah M, Gerlach JQ, Bhavanandan V, Nagaraj V, Smith AD, Fujiyama K, Sommer U, Costello CE, Olszewski N, Joshi L (2009). O-glycosylation of protein subpopulations in alcohol-extracted rice proteins.
J Plant Physiol166,219–232.
Kimura Y, Watanabe T, Kimura M, Maeda M, Murata Y, Fujjyama K (2008). Salt-adaptation of tobacco BY2 cells induces change in glycoform of N-glycans: enhancement of exo- and endo-glycosidase activities by salt-adaptation.
Biosci Biotechnol Biochem 72,514–522.
Kishimoto T, Watanabe M, Mitsui T, Hori H (1999).
Glutelin basic subunits have a mammalian mucin-type O-linked disaccharide side chain. Arch Biochem Biophys 370,271–277.
Laus MC, Logman TJ, Lamers GE, van Brussel AAN, Carlson RW, Kijne JW (2006). A novel polar surface polysaccharide from Rhizobium leguminosarum binds
528 植物学报 45(5) 2010
host plant lectin. Mol Microbiol59,1704–1713.
Ohto M, Onai K, Furukawa Y, Aoki E, Araki T, Nakamura K (2001). Effects of sugar on vegetative development and floral transition in Arabidopsis. Plant Physiol127, 252–261.
Rademacher TW, Parekh RB, Dwek RA (1988).
Glycobiology. Annu Rev Biochem57,785–838.
Riou-Khamlichi C, Menges M, Healy JMS, Murray JAH (2000). Sugar control of the plant cell cycle: differential regulation of Arabidopsis D-type cyclin gene expression.
Mol Cell Biol20,4513–4521.
Rolland F, Baena-Gonzalez E, Sheen J (2006). Sugar sensing and signaling in plants: conserved and novel mechanisms. Annu Rev Plant Biol57,675–709.
Rudiger H, Gabius HJ (2001). Plant lectins: occurrence, biochemistry, functions and applications. Glycoconj J18, 589–613.
Saint-Jore-Dupas C, Nebenfuhr A, Boulaflous A, Follet-Gueye ML, Plasson C, Hawes C, Driouich A, Faye L, Gomord V (2006). Plant N-glycan processing enzymes employ different targeting mechanisms for their spatial arrangement along the secretory pathway. Plant Cell18,3182–3200.
Seveno M, Cabrera G, Triguero A, Burel C, Leprince J, Rihouey C, Vezina LP, D'Aoust MA, Rudd PM, Royle L, Dwek RA, Harvey DJ, Lerouge P, Cremata JA, Bardor M (2008). Plant N-glycan profiling of minute amounts of material. Anal Biochem379,66–72.
Shimada A, Ueguchi-Tanaka M, Sakamoto T, Fujioka S, Takatsuto S, Yoshida S, Sazuka T, Ashikari M, Matsuoka M (2006). The rice SPINDLY gene functions as
a negative regulator of gibberellin signaling by controlling
the suppressive function of the DELLA protein, SLR1, and modulating brassinosteroid synthesis. Plant J48, 390–402.
Shridhar S, Chattopadhyay D, Yadav G (2009). PLecDom:
a program for identification and analysis of plant lectin
domains. Nucleic Acids Res37,W452–W458.
Strasser R, Altmann F, Glossl JGR, Steinkellner H (2004). Unaltered complex N-glycan profiles in Nicotiana benthamiana despite drastic reduction of beta 1,2-N-acetylglucosaminyltransferase I activity. Glycoconj J21,275–282.
Strasser R, Mucha J, Schwihla H, Altmann F, Glossl J, Steinkellner H (1999). Molecular cloning and characterization of cDNA coding for beta 1,2-N-acetylglu- cosaminyltransferase I (GlcNAc-TI) from Nicotiana tabacum. Glycobiology9,779–785.
Vandenborre G, van Damme EJM, Smagghe G (2009).
Natural products: plant lectins as important tools in controlling pest insects. In: Ishaaya I, Horowitz AR, eds.
Biorational Control of Arthropod Pests: Application and Resistance Management. Dordrecht: Springer-Verlag.
pp.163–187.
Wang XQ (2009). Structure, mechanism and engineering of plant natural product glycosyltransferases. FEBS Lett 583,3303–3309.
Wu AM, Lisowska E, Duk M, Yang Z (2009). Lectins as tools in glycoconjugate research. Glycoconj J 26, 899–913.
Xing LJ, Li J, Xu YY, Xu ZH, Chong K (2009).
Phosphorylation modification of wheat lectin VER2 is associated with vernalization-induced O-GlcNAc signaling and intracellular motility. PLoS One3, e4854.
doi:10.1371/journal.pone.0004854.
Yang SM, Chen ZC, Jin X, Lin XF (2006). HRP biosensor based on sugar-lectin biospecific interactions for the determination of phenolic compounds. Electrochim Acta 52,200–205.
Ye SH, Chen S, Zhang F, Wang W, Tian Q, Liu JZ, Chen F, Bao JK (2009). Transgenic tobacco expressing zephyranthes grandiflora agglutinin confers enhanced resistance to aphids. Appl Biochem Biotechnol158, 615–630.
尹恒等:植物糖生物学研究进展 529 Research Progress in Plant Glycobiology
Heng Yin, Wenxia Wang, Xiaoming Zhao*, Yuguang Du*
Liaoning Provincial Key Laboratory of Carbohydrates, Dalian Institute of Chemical Physics, Chinese Academy of Sciences,
Dalian 116023, China
Abstract Abundant results on glycobiology were achieved in animal and microbe research fields after the glycobiology theory was proposed in 1988, but little progress was made in plants. Furthermore, few systematic monographs or reviews have focused on plant glycobiology. In this paper, we promote the concept of plant glycobiology, which concentrates on the plant-saccharide interaction mechanism, as well as the structure and biology function of saccharides (sugar chains or glycans). We review our results and those of others, especially the latest progress, on several aspects of plant glycobi-ology such as sugar signaling under normal development and stress conditions, plant glycoproteins and glycans, and plant glycosyltransferases and lectins.
Key words glycoprotein, glycosyltransferase, lectin, plant glycobiology, sugar signaling
Yin H, Wang WX, Zhao XM, Du YG (2010). Research progress in plant glycobiology. Chin Bull Bot45, 521–529.———————————————
*Author for correspondence. E-mail: zxm@https://www.360docs.net/doc/4c3467283.html,; dyguang@https://www.360docs.net/doc/4c3467283.html,
(责任编辑: 刘慧君)
考研农学联考植物生理学真题参考复习资料
2011 年考研农学联考植物生理学真题参考答案 一、单项选择题:I?15小题,每小题1分,共15分。下列每题给出的四个选项中,只有一个选项是符合题目要求的。 1. G- 蛋白是一类具有重要生理调节功能的蛋白质,它在细胞信号转导中的作用是 A. 作为细胞质膜上的受体感受胞外信号 B. 经胞受体激活后完成信号的跨膜转换 C. 作为第二信号 D. 作为蛋白激酶磷酸化靶蛋白 【参考答案】B 【考查知识点】植物细胞信号转导一GTP结合调节蛋白作用 2. 植物细胞进行无氧呼吸时 A. 总是有能量释放,但不一定有C02释放 B. 总是有能量和C02释放 C. 总是有能量释放,但不形成ATP D. 产生酒精或乳酸,但无能量释放 【参考答案】A 【考查知识点】植物呼吸代谢及能量转换—无氧呼吸特点 3. 以下关于植物细胞离子通道的描述,错误的是 A. 离子通道是由跨膜蛋白质构成的
B. 离子通道是由外在蛋白质构成的 C. 离子通道的运输具有一定的选择性 D. 离子通道的运输只能顺电化学势梯度进行 【参考答案】B 【考查知识点】植物细胞跨膜离子运输一离子通道的特点 4. C3植物中,RuBp竣化酶催化的C02固定反应发生的部位是 A. 叶肉细胞基质 B. 叶肉细胞叶绿体 C. 维管束鞘细胞机制 D. 维管束鞘细胞叶绿体 【参考答案】B 【考查知识点】光合作用一RuBP竣化酶催化部位 5. 细胞壁果胶质水解的产物主要是 A. 半乳糖醛酸 B.葡萄糖 C.核糖 D.果糖 【参考答案】A 【考查知识点】细胞壁一细胞壁的果胶质水解产物 6. 叶片衰老过程中最先解体的细胞器是 A. 高尔基体 B.内质网 C.叶绿体 D.线粒体
《植物学》期末考试试题及参考答案 (4)
生物科学专业 植物学期末考试参考答案 姓名_______专业与班级_________学号__________成绩________ 我保证诚实地完成本次考试,保证不以任何形式抄袭或作弊。 自愿签约人: 一.填空题(除注明者外,每空0.5分,共40分) 1.(3分)裸子植物苏铁纲具有羽状分裂大孢子叶,银杏纲的大孢子叶称为珠领, 松柏纲的大孢子叶称为珠鳞,紫杉纲的大孢子叶称为珠托或套被,而买麻藤纲具有盖被和珠孔管。 2.(2.5分)犁头尖具佛焰花序,花序顶端为附属体,依次以下为雄花 花,中性花花,雌花花。 3.(3分)芸香科植物的识别特征有:托叶缺,枝叶树皮香气,叶具透明 油腺点,花盘存在于雄蕊与子房之间;果实有多种,柑桔属的果实称为柑果,花椒属的果实称为蓇葖。 4.(2.5分)大戟科叶具有托叶、有些植物具乳汁、叶柄上端或叶上常有腺体、花 单性、三室果。 5.(3分)樟科的识别特征有枝叶树皮有香气,花三数,雄蕊四轮时,每轮3枚, 第1与第2轮雄蕊花药内向,第3轮雄蕊花药外向,基部有2枚腺体,第4轮雄蕊退化为箭头状。花药瓣裂。 6.(2.5分)填空: (1)黄鹌菜的头状花序上的花全为舌状花 (2)板栗的果实由壳斗包 (3)草莓食用部是花托 (4)柚的叶是单身复叶
(5)咖啡属于茜草科。 7.(1.5 分)山毛榉科植物的雄花序是柔荑花序;杨柳科花单性,无花被,杨属 的花具有花盘,而柳属的花具有蜜腺。 8.(3分)构成禾本科植物花序的基本单位称小穗, 其基部两侧各有一枚颖片, 每 一两性小花由外稃、内稃、2~3枚浆片、6~3枚雄蕊和雌蕊组成。 9.水杉具有条形且对生的叶。 10.(4.5分)木兰科的原始特征包括花各部多数,分离,螺旋状排列在伸 长的花托上,雄蕊花药长,花丝短,药隔突出;雌蕊花柱与柱头不分化。 11.(2分)具有双韧维管束的植物类群有葫芦科、桃金娘科、夹竹桃科、萝 摩科等。 12.(2分)桃金娘科叶对生,具边脉,常有香气和油腺点,子房半下位。 13.(2.5分)有些植物可根据营养器官的特征来判别所属的科, 例如茎节膨大,叶 常互生,具有鞘状或穿茎托叶的是蓼科;植物体有香气,托叶脱落后在枝条上留下托叶环的是木兰科;有些植物可根据繁殖器官特征来判别所属的科,例如具双悬果的是伞形花科;具角果的是十字花科;具有瓠果的是葫芦科。 14.写出下列植物的学名(每小题1分,共5分): (1)银杏:Ginkgo biloba。 (2)月季:Rosa chinensis。 (3)大豆:Glycine max。 (4)水稻:Oryza sativa。 (5)无花果属:Ficus。 15.(3分)锦葵科植物最易识别的特征是单体雄蕊;苋科与藜科均具有基生 胎座,前者的花被常有颜色,后者的花被绿色。石竹科石竹科花瓣反折,前端流苏状撕裂,有爪。 二.选择题(把选择答案前面的字母直接写在该题的双括号内,每小 题1分,共5分)
植物糖生物学研究进展
植物学报 Chinese Bulletin of Botany 2010, 45 (5): 521–529, https://www.360docs.net/doc/4c3467283.html, doi: 10.3969/j.issn.1674-3466.2010.05.001 —————————————————— 收稿日期: 2010-01-18; 接受日期: 2010-03-23 基金项目: 863计划(No.2006AA10A213, No.2007AA091601)和中国科学院知识创新工程重要方向项目(No. KSCX2-YW-G-041) * 通讯作者。E-mail: zxm@https://www.360docs.net/doc/4c3467283.html,; dyguang@https://www.360docs.net/doc/4c3467283.html, 植物糖生物学研究进展 尹恒, 王文霞, 赵小明*, 杜昱光* 中国科学院大连化学物理研究所辽宁省碳水化合物重点实验室, 大连 116023 摘要 自1988年糖生物学概念提出以来, 国内外科学家在动物、微生物领域取得了大量的研究成果, 但植物糖生物学的研究进展较慢, 目前少见系统的专著或综述。该文围绕植物正常生长时糖信号、逆境时糖信号、糖蛋白及其糖链、重要糖基转移酶及植物凝集素等植物糖生物学的主要问题, 全面阐述植物糖生物学的各个研究分支, 并介绍各领域的最新研究进展。提出了植物糖生物学的概念, 并将其定义为研究植物与糖类互作机制及植物体内糖(糖链与糖分子)结构及生物学功能的科学。 关键词 糖蛋白, 糖基转移酶, 凝集素, 植物糖生物学, 糖信号 尹恒, 王文霞, 赵小明, 杜昱光 (2010). 植物糖生物学研究进展. 植物学报 45, 521–529. 糖类是生物体的重要组成成分, 在自然界中分布广泛, 含量丰富。但直到20世纪上半叶, 糖类仍被视为是缺乏生物特异性的一类惰性化合物, 只是作为代谢能量来源或充当结构保护材料(如植物细胞壁和昆虫的外壳), 在生物体内功能较少。由于糖类物质结构复杂、糖链分析技术缺乏, 科学家们对其研究关注不多, 使得糖类的研究远远落后于另2种生物大分子 ——核酸和蛋白质。 20世纪70年代以来, 随着糖链解析技术水平的提高以及分子生物学的发展, 尤其是人、拟南芥(Arabidopsis thaliana )等模式生物基因组测序的完成, 围绕糖类物质的研究工作日渐增多。越来越多的证据表明, 糖类物质全面参与了生物的生殖发育、生长、应激等过程, 是很多生理和病理过程中分子识别的决定因素。最初, 这些围绕糖的研究工作被认为是糖化学的一个分支, 但很快其中大量的生物学工作远远超出了糖化学的范畴, 因此科学家们提出了糖生物化学的概念, 而随着研究内容的进一步深入, 糖生物化学也不能完全涵盖糖在生物领域的最新研究进展。1988年, 生化领域的著名杂志《生物化学年评》发表了英国牛津大学Rademacher 等人题为“糖生物学(Glycobiology)”的一篇综述文章(Rademacher et al., 1988), 标志着糖生物学这一学科的正式诞生。此后, 围绕着糖链结构及糖的生物学功能, 科学家们在糖链与疾病的关系、天然产物中糖的分离提纯以及功能糖的制备与应用等方面进行了大量的工作, 取得了一定进展。2001年, Science 杂志汇编了Hurtley 等人的7篇综述和6篇简介, 以《灰姑娘的马车来了》为题编辑了一期“糖和糖生物学”专辑, 对糖生物学最新的研究成果及前景进行了综述和展望, 从而将糖生物学的研究推向了一个新的高度(Hurtley et al., 2001)。2006年, Nature 杂志也推出了糖化学与糖生物学的专辑, 全面介绍了糖生物学领域的研究进展。我国糖生物学的开展与国际接轨较快, 1995年金城等人将糖生物学概念引入中国(金城和张树政, 1995), 此后, 我国科学家在糖生物合成和糖链功能解析等领域取得了一定进展。 广义糖生物学的含义是: 研究自然界中广泛分布的糖(糖链或聚糖)的结构、生物合成和生物学意义。但有关糖类结构和生物合成的研究也是已有学科糖化学和糖生物化学的主要研究内容之一, 所以糖生物学研究和讨论的对象更多地聚焦在一些重要的功能糖、生物体内糖缀合物的生物学功能上。实际上, 糖生物学的研究焦点是糖类和其它分子的关系, 有一种观点认为, 蛋白质和糖类的相互作用是糖生物学的基础(王克夷, 2009)。目前糖生物学的工作多围绕动物、 ·特邀综述·
植物生物学试题及答案
植物学试题 名词解释(12分,2分/个) 1、细胞器; 2、木质部脊; 3、束中形成层; 4、完全叶; 5、花程式; 6、聚合果 二、判断与改错(17分,对的填“ +”,错的填“-”并将错的改正) 1、质体是一类与碳水化合物合成及贮藏相关的细胞器。() 2、根毛分布在根尖的伸长区和成熟区。() 3、根的初生木质部发育顺序为外始式,而在茎中则为内始式。() 4、水生植物叶小而厚,多茸毛,叶的表皮细胞厚,角质层也发达。() 5、胡萝卜是变态根,主要食用其次生韧皮部。() 6、观察茎的初生结构和次生结构都是从节处取横切面。() 7、由3心皮组成的复雌蕊有6条腹缝线。() 8、二体雄蕊就是一朵花中只有两个离生的雄蕊。() 9、双受精是种子植物的重要特征之一。() 10、桃和梨均为假果。() 三、填空(21分,0.5分/空) 1、植物细胞的基本结构包括 _和_两大部分。后者又可分为___________ 、____ 和___ 三部分。 2、保护组织因其来源及形态结丽不厂可分为___________ 和___ 。 3、兰麻等纤维作物所利用的是组织。 4、植物种子是由 , 和三部分构成,但有些种子却只有和两部分,前者称种子,后者 称种子。 5、禾物的内皮层在发育后期其细胞常面增厚,横切面上,增厚的部分呈形。 6、双子叶植物和裸 子植物茎的初生结构均包括、和三部分,玉米等单子叶植 物茎的初生结构包括、和三部分。 7、双子叶植物茎的维管束多为,在初生木质部和初生韧皮部间存在形成层,故又称维管束。 8、叶的主要生理功能包括和等两个方面。 9、一朵完整的花可分为____ 、____ 、___ 、_____ 、 ___ 和____ 六部分。其中最主要的部分是_______ 和_____ 。 10、小抱子母细胞进行减^^前?1雨101二般岳厂、、和组成。花粉成熟时,花粉囊壁一般 只留下和。 11、被子植物细胞的减数分裂在植物的生活史中发生次。 四、选择题(10分,1分/个) 1、光学显微镜下呈现出的细胞结构称。 A .显微结构;B.亚显微结构;C.超显微结构;D.亚细胞结构 2、裸子植物靠输导水分。 A .导管和管胞;B.管胞;C.筛管;D .筛胞 3、成熟蚕豆种子的种皮上一条黑色眉状物是。 A .种脊;B.种脐;C.种阜;D.种柄 4、原表皮、基本分生组织和原形成层属于。 A .居间分生组织;B.原分生组织;C.初生分生组织; D.伸长区 5、在方形(如蚕豆)或多棱形(如芹菜)的茎中,棱角部分常分布有。 A .厚角组织;B.厚壁组织;C.薄壁组织;D.石细胞 6、栅栏组织属于。 A .薄壁组织;B.分生组织;C.保护组织;D.机械组织 7、以下所列的结构,哪一些都是茎的变态?。 A .皂荚的分枝刺,葡萄和豌豆的卷须; B.马铃薯、姜、摹养、芋头; C .莲藕、菊芋、竹鞭、白萝卜、胡萝卜; D .蔷薇和刺槐(洋槐)的刺,豌豆的卷须 8、具的花为整齐花。 A .舌状花冠;B.唇形花冠;C.蝶形花冠;D.十字花冠
1糖生物学
1糖生物学 科学家把研究生物体内多糖的科学叫做“糖生物学”,也有人沿袭“基因组学”和“蛋白质组学”的概念把这们学科叫做“糖原组学”。糖生物学这一个名词的提出是在1988年。牛津大学德威克教授在当年的《生化年评》中撰写了以“糖生物学”为题的综述,这标志了糖生物学这一新的分支学科的诞生。 研究对象 糖生物学(glycobiology)是研究聚糖及其衍生物的结构,化学,生物合成及生物功能的科学 蛋白质、核酸和多糖是构成生命的三类大分子,蛋白质和核酸的研究已经成为生命科学中的热点问题。糖类的研究一度被人遗忘,只有少数科学家在苦苦探索着糖类的奥秘,糖类研究成了生命科学中的灰姑娘。然而,随着蛋白质和核酸(主要是基因的研究)中更多的奥秘被人类知晓,糖类的重要性也浮出水面,成为了医学研究的“甜蜜之点”,糖类研究这个“灰姑娘”等来了属于她自己的马车。科学家认为,糖类的研究将像一个人见人爱的“甜苹果”一样,获得更多科学家的青睐,将成为生命科学研究中的新热点。 2糖生物学的崛起 科学家把研究生物体内多糖的科学叫做“糖生物学”,也有人沿袭“基因组学”和“蛋白质组学”的概念把这们学科叫做“糖原组学”。糖生物学这一个名词的提出是在1988年。牛津大学德威克教授在当年的《生化年评》中撰写了以“糖生物学”为题的综述,这标志了糖生物学这一新的分支学科的诞生。[1]同一年牛津大学研制成功了N-糖链的结构分析仪,而且将它商品化。 将糖生物学推向生命科学前沿的重大事件发生于1990年。有3家实验室几乎同时发现血管内皮细胞-白血球粘附分子1(ELAM-1),后来改名为E-选凝素 (E-selectin)。这一位于内皮细胞表面的分子能识别白血球表面的四糖 Sia-LeX。当组织受到损伤时,白血球和内皮细胞穿过血管壁,进入受损组织,以便杀灭入侵的异物。然而,过多白血球的进入则可能导致炎症的产生。这一发现首次阐明了炎症过程有糖类和相关的糖结合蛋白参与。更令人吃惊的是,进入血液循环系统的癌细胞可能借助了类似于上述的机制穿过血管,进而导致癌症的转移。紧接着又出现了以这一基础研究的成果为依据的开发和生产抗炎和抗肿瘤药物的热潮。[1] 3攻克疾病的“甜苹果”
植物生物学实验教案—优秀教案
植物生物学实验教案授课专业:生物科学、农学主讲:
实验1 光学显微镜及体视镜的构造和使用方法 一、实验目的 1、了解光学显微镜及体视镜的一般构造和性能; 2、学会正确地使用光学显微镜及体视镜,熟练地掌握对光,低高倍物镜的使用技术, 以及显微镜的维护; 3、学会临时装片的制作和徒手切片。 二、重点与难点 正确地使用光学显微镜及体视镜,熟练地掌握对光,低高倍物镜的使用技术。三、教学方法与手段 本次课主要采取讲授法和讨论法,在学生实验过程中辅以个别指导进行教学。 四、实验内容 1、光学显微镜的构造、使用方法及维护; 2、临时装片的制作及徒手切片的练习; 3、体视镜的一般结构及使用方法。 五、实验材料 洋葱(Allium cepa)根尖永久装片;洋葱(Allium cepa)鳞片叶;油菜(Brassica campestris)或水稻(Oryza sativa)花粉。 六、实验用品 普通光学显微镜、体视显微镜;镊子、载玻片、盖玻片、培养皿、纱布、吸水纸、擦镜纸、滴瓶、毛笔;碘液、水。 七、实验方法 (一)普通光学显微镜的构造、使用方法及维护 1、显微镜的构造 显微镜的基本结构可以分两部分,即光学部分与机械部分。 (1)光学部分 ①物镜、②目镜、③聚光器、④虹彩光圈、⑤反光镜、⑥镜筒 (2)机械部分 ①镜座、②镜柱、③镜臂、④载物台、⑤物镜转换器、⑥调焦螺旋 2、显微镜的使用方法 (1)正确安置显微镜、(2)对光、(3)低倍物镜的使用、(4)高倍物镜的使用(5)浸油物镜的使用、(6)显微镜的使用练习、(7)用毕复原 3、显微镜的放大倍数 4、光学显微镜的显微测微法 (1)显微测微计 ①镜台测微计、②目镜测微计 (2)测量方法
植物生物学试题及答案
植物学试题一名词解释(12分,2分/个) 1、细胞器; 2、木质部脊; 3、束中形成层; 4、完全叶; 5、花程式; 6、 聚合果 二、判断与改错(17分,对的填“+”,错的填“-”并将错的改正) 1、质体是一类与碳水化合物合成及贮藏相关的细胞器。( ) 2、根毛分布在根尖的伸长区和成熟区。( ) 3、根的初生木质部发育顺序为外始式,而在茎中则为内始式。( ) 4、水生植物叶小而厚,多茸毛,叶的表皮细胞厚,角质层也发达。( ) 5、胡萝卜是变态根,主要食用其次生韧皮部。( ) 6、观察茎的初生结构和次生结构都是从节处取横切面。( ) 7、由3心皮组成的复雌蕊有6条腹缝线。( ) 8、二体雄蕊就是一朵花中只有两个离生的雄蕊。( ) 9、双受精是种子植物的重要特征之一。() 10、桃和梨均为假果。()
三、填空(21分,分/空) 1、植物细胞的基本结构包括和两大部分。后者又可分为、和三 部分。 2、保护组织因其来源及形态结构的不同,可分为和。 3、苎麻等纤维作物所利用的是组织。 4、植物种子是由,和三部分构成,但有些种子却只有和两部 分,前者称种子,后者称种子。 5、禾本科植物的内皮层在发育后期其细胞常面增厚,横切面上,增厚的部分呈形。 6、双子叶植物和裸子植物茎的初生结构均包括、和三部分,玉米等单子叶植 物茎的初生结构包括、和三部分。 7、双子叶植物茎的维管束多为,在初生木质部和初生韧皮部间存在形成层,故 又称维管束。 8、叶的主要生理功能包括和等两个方面。 9、一朵完整的花可分为、、、、和六部分。其中最主 要的部分是和。 10、小孢子母细胞进行减数分裂前,花粉囊壁一般由、、和组成。
植物生物学复习题
0绪论复习题 1.什么是植物?在林奈的二界系统和魏泰克的五界系统中,植物包括的范围有何变化? 植物有明显的细胞壁和细胞核,其细胞壁由纤维素构成,具有光合作用的能力——就是说它 可以借助光能及动物体内所不具备的叶绿素,利用水、矿物质和二氧化碳生产食物。魏泰克的五界系统中不仅包括林奈的二界系统中的植物界和动物界,还增加了真菌界,原生生物界,原核生物界。 2.列举5个我国著名的植物研究机构,简述他们的主要研究领域。 ○1中国科学院植物研究所(系统与进化植物学领域、植物生态学(草原)、光合作用、植物分子生理与发育领域等);○2中国科学院昆明植物研究所(植物分类与生物地理、植物化学 与天然产物研发、野生种质资源保藏与利用、民族植物学与区域发展、资源植物研发与产业化);○3中国农业大学,主要研究领域:植物逆境机理、植物发育生物学、作物重要性状功 能基因组学、植物基因表达调控的分子机理;○4中国科学院上海生命科学研究院植物生理生 态研究所(功能基因组学,分子生理与生物化学,环境生物学和分子生态学等);○5中国科学院上海植物逆境生物学研究中心(植物逆境分子生物学研究)。 3.列举5个我国当代著名的植物学家,简述他们的主要研究领域。 张新时院士,植物生态学;洪德元院士,植物细胞分类学;王文采院士,植物分类学;匡廷 云院士,光合作用;周俊院士,植物化学;施教耐院士,植物呼吸代谢;陈晓亚院士,植物 次生代谢。 01细胞与组织-01细胞-复习题 一、选择 1.光镜下可看到的细胞器是。 A.微丝B.核糖体C.叶绿体D.内质网 2.光学显微镜下呈现出的细胞结构称。 A.显微结构B.亚显微结构C.超显微结构D.亚细胞结构 3.下列细胞结构中,具单层膜结构的有。 A.叶绿体B.线粒体C.溶酶体D.核膜E.液泡 4.下列细胞结构中,具双层膜结构的有, A.叶绿体B.线粒体C.溶酶体G.微管I.高尔基体J.内质网K.核膜 5.植物细胞初生壁的主要成分是。 A.纤维素、半纤维素和果胶B.木质、纤维素和半纤维素C.果胶D.角质和纤维素 6.初生纹孔场存在于。 A.次生壁B.初生壁C.胞间层D.角质层 7.糊粉粒贮藏的养分是。 A.淀粉B.脂肪C.蛋白质D.核酸 8.细胞进行呼吸作用的场所是。 A.线粒体B.叶绿体C.核糖体D.高尔基体 9.与细胞分泌功能有关的细胞器是。 A.线粒体B.高尔基体C.溶酶体D.白色体 10.细胞内有细胞活动的“控制中心”之称的是。 A.细胞器B.细胞核C.细胞质D.叶绿体
植物生物学试题
被子植物 1.合蕊柱 2.叶枕 3.距 4.冠毛 5.蝶形花冠和假蝶形花冠 二、填空题 1. Cathaya argyrophyllum中名是_____属_____科 Magnolia denudate是_____科植物Cercis chinensis中名是_____,是_____科(亚科)植物 Panax ginshen中名是_____,是_____科植物 Rhododendron smithii中名是_____,是_____科植物 Lonicera japonica中名是_____,是_____科植物 Allium cepa中名是_____,是是_____科植物。 2. _____ 科的特征是木本,单叶互生;雄蕊、雌蕊多数,螺旋状排列在伸长的花托上,聚合蓇葖果。 3. _____ 科的特征是头状花序,聚药雄蕊,果具冠毛。 4. _____ 科的特征是雄花成柔荑花序,雌花生于总苞中。 5. _____ 科的特征是有副萼,单体雄蕊,蒴果。 6. _____ 科的特征是十字花冠,四强雄蕊,角果。 7. _____ 科的特征是有托叶,叶基有腺体,周位花,核果。 8. _____ 科的特征是蝶形花冠,荚果。 9. _____ 科的特征是具乳汁,花单性,子房 3 室,中轴胎座。 科的特征是蔓生草本,有卷序,瓠果。 科的特征是叶具叶舌,颖果。 科的特征是具唇瓣,合蕊柱。 13.百合科的花程式是 _____。 14.假花学说认为被子植物起源于 _____ 植物,因而 _____ 类是最原始的代表。 15.真花学说认为每一个雄蕊代表 _____ ,心皮是 _____ 变化而来的,因而 _____ 类植物是最原始的,这一学说得到了广泛的承认,同时由于被子植物具有 _____、_____、_____ 三个特点,被认为是由共同起源单元说。 16.下列植物各属于哪个科。 17.矮牵牛属 ______ 科;金鱼草属 ______ 科;辛夷属 ______ 科;吊兰属 ______ 科。胡萝卜 _____ 科;马铃薯 _____ 科;白菜 _____ 科;番茄 _____ 科;香蕉 _____ 科;
植物生物学参考试题及解析
植物生物学参考试题及解析 LYT&FLN 一、名词解释: 原生质:细胞内有生命活性的物质,组成原生质体,物理性质是一种半透明的亲水胶体。 原生质体:是指单个细胞内的分化了原生质,是细胞的最主要部分,细胞的一切代谢活动都在这里进行。 显微结构:光学显微镜下观察到的结构,由细胞壁和原生质体构成。 亚显微结构:在电子显微镜下显示的细胞结构,包括细胞器结构。 不活动中心:处于植物根尖顶端分生组织中心部分组成一个区域,不常分裂,大小变化很小,合 成核酸和蛋白质的速率很低。 内皮层:皮层最内方的一层细胞,细胞小、排列紧密,各细胞的径向壁和上下横壁有带状的木化和栓化加厚区域,称为凯氏带。 髓射线:中柱内的薄壁组织,是由原形成层以内的基本分生组织分化而来。 补充组织:周皮形成过程中,在原来气孔位置下面的木栓形成层不形成木栓细胞,而产生一团圆球形,排列疏松的薄壁细胞 叶痕:茎上的叶片脱落后留下的疤痕 叶脉:贯穿在叶肉内的维管组织及外围的机械组织。由分布在叶片中的维管束及其周围的有关组织组成,起支持和输导作用。 完全花:具有萼片、花瓣、雄蕊、雌蕊的花。 不完全花:缺少萼片、花瓣、雄蕊、雌蕊中的一种或两种的花。 开花:当雄蕊中的花粉粒和雌蕊中的胚囊或二者之一已经成熟。花冠,花萼张开,雌、雄蕊露出的现象。 传粉:花被打开,花药开裂,成熟的花粉粒借外力传送到雌蕊柱头上的过程。 双名法:属名+种名+定名人的姓氏或姓名缩写. 生物多样性:指地球上的生物及其与环境形成的所有形式、层次和联合体的多样化。 共质体:通过胞间连丝结合在一起的原生质体。 质外体:共质体以外的部分。 胞间层:又称中胶层,是细胞分裂产生新细胞时形成的,由相邻的两个细胞向外分泌果胶物质构成共有的一层薄膜。初生壁:是细胞增长体积时所形成的壁层,由相邻细胞分别在胞间层两面沉积壁物质而成。 二、概念: 细胞壁以内的溶液是什么?(答案:原生质体,其中的溶液为细胞质基质和细胞液) 成熟的导管分子和筛管分子都是死细胞么?为什么?(答案:成熟的导管分子为长管状的死细胞, 无生活的原生质体,成熟的筛管为长形活细胞,筛管分子长成后,细胞核退化,但细胞质仍保留,与有机 物的运输有关。) G0期细胞还能分化么?为什么?(提示:能,G0细胞在形成之后不再进行DNA的复制,暂时脱离细 胞周期的细胞(休止期),适当刺激即可重新进入细胞周期G1期,恢复分裂、分化能力,进行增殖) 维管植物的主要组织可归纳为哪三个系统?(答案:、维管系统、基本组织系统、皮系统) 主根和侧根均属于初生根么?(答案:不是) 初生木质部和初生韧皮部各三束,应该是几原型?(答案:三原型) 次生结构中仍有皮层存在么?(答案:没有,由周皮代替外皮层起保护作用) 不定根发生在哪里?(答案:中枢鞘) 等面叶的主要代表物种有哪些?(答案:禾本科水稻) 通气组织是什么?(答案:水生或湿生植物体中,由发达的胞间隙,形成的气腔或气道。贮存或交换气 体,抵抗机械应力。)
普通生物学实验讲义
普通生物学实验(动物、植物) 实验一(1)普通光学显微镜及其使用 一、实验目的 了解普通光学显微镜的构造及其原理,并熟练掌握其操作方法。 二、实验用品 普通复式光学显微镜、载玻片、盖玻片、滤纸、擦镜纸。 三、实验原理和方法 普通光学显微镜从构造上可分光学、机械和电子三大系统。 1、显微镜的光学系统 光学系统通常由物镜、目镜、聚光器和光阑组成。 1)、物镜(objective) 显微镜的质量主要取决于物镜。物镜种类繁多,性能相差悬殊。物镜的放大倍数用数字表示,如4、10、20、40和100等。 a.干燥系(drysystem)物镜 镜检时,物镜与盖片间,不添加任何液体。如4×、10×、20×和40×物镜都属干燥系,使用时不加用任何浸液,只以空气为介质,其折射率为1,所以干燥系物镜的数值孔径小,分辨率亦低。 b.浸没系(immersionsystem)物镜 物镜在使用时,前透镜与盖片之间浸满液体。依充添的浸液的不同,主要可分为油浸系(oil immersion)和水浸系(water immersion)等类别。最常用的浸没液为香柏油(cederoil),其折射率为1.515,与玻片的折射率相近,且不易干涸。使用水浸物镜时加用水,其折射率1.33。 油浸物镜外壳上刻有:“oil”、“oel”、“imm”和“HI”等字样,水浸物镜刻有“W”或“Water”字样;油、水浸两用物镜则刻上“oil+w”字样;甘油浸没物镜刻有“Glyc”或“Glyz”等字样。 2)、目镜(eyepiece,ocular) 目镜作为影像和肉眼间的放大镜,将物镜映来的影像做第二次放大。同时,目镜作为物镜的补偿,把物镜残留下的像差给予进一步校正,以提高造像质量。目镜作为投影器,把放大的影像投射在摄影暗箱的焦平面上。 目镜通常由两片(组)正透镜组成,上面的透镜叫接目或眼透镜(eye—lens),它决定倍数和成像的优劣;下面的透镜叫会聚透镜(collectivelens)或场镜(fieldpiece),它使视野边缘的成像光线向内折射,进入眼透镜中,使物体的影像均匀明亮。上下透镜的中点,或场镜下面设有用金属制造的光阑叫做视野光阑或场光阑(field stop)。场镜或物镜在这个光阑面造像,在光阑上可装入各种目镜测微计、十字线玻片和指针等。由眼透镜射出的成像光线基本上为平行光束,并在目镜之上约10mm处交叉,此交叉点称作出射光瞳。 3)聚光器(condensers)
最新植物生理学研究生考试题及答案
植物生理学2015年研究生考试题及答案 一、填空题(每空1分,共计28分) 1、海芋植物的佛焰花序比一般植物的呼吸放出的热量比一般植物高,是因 为存在抗氧呼吸的缘故。 2、与植物耐旱性有重要相关性的氨基酸是,它能增强细胞 的。 3、植物叶绿体的丙酮提取液透射光下呈,反射光下 呈。 4、根据种子的吸水量,可将种子的萌发分为吸胀吸水阶段、停止吸水阶段,重 新吸水阶段。 5、GA和ABA生物合成的前体是甲瓦龙酸,在短光照下形成ABA。 6、膜脂的组成与膜脂的抗冷性有关,不饱和程度,固化温度 高,不利发生膜变相,植物的抗冷性越小。 7、植物组织培养的理论基础是细胞全能性,用来培养的植物体部分叫外植 体。 8、保卫细胞质的膜上存在着 H+ATP 酶,在光照下,将H+分泌到保卫细胞外, 使保卫细胞 HP升高,驱动 H+ 进入保卫细胞,导致保卫细胞吸水,气孔张开。 9、跨膜信号传导主要是通过和完成。 10、土壤缺氮时,根冠比高,水分过多时,根冠比低。 11、具有远红光和红光逆转效应的是,它的生色团与叶绿体 的 结构相似。 12、成熟的水果变甜,是因为淀粉转化成糖,未成熟的水果有涩味是因为 含有单宁。 13、植物组织培养的理论依据是细胞全能性,用来培养的植物的部分叫外 植体。 二、单项选择(每题1分,共计20分) 略!
三、名词解释(每题3分,共计30分) 1、次级共运转(次级主动运输):以质子动力作为驱动力的跨膜离子运转,使质 膜两边的渗透能增加,该渗透能是离子或者中性分子跨膜转运的动力。 2、细胞信号传导:偶联各种胞外刺激信号与其相应的生理反应之间的一系列分 子反应。 3、希尔反应:离体叶绿体在光下所进行的分解水并放出氧气的反应。 4、渗透调节:植物细胞通过主动增加溶质降低渗透势,增强吸水和保水能力, 以维持正常细胞膨压的作用。 5、交叉适应:植物经历了某种逆境之后,能提高对另一逆境的抵抗能力,对不 同逆境间的相互适应作用。 6、光饱和点:在一定范围内,光合速率随着光照强度的增加而加快,光合速率 不再继续增加是的光照强度称为光饱和点。 7、光的形态建成:依赖光控制细胞分化、结构和功能的改变,最终汇集成组织 和器官的建成,就称为光形态建成。 8、极性运输:生长素只能从植物体形态学上端向下端运输,不能反之。 9、单盐毒害:植物培养在单盐溶液中所引起的毒害作用. 10、水孔蛋白:存在于生物膜上的一类具有选择性、高效转运水分功能的内 在蛋白。 四、简答题(每题7分,共计42分) 1、生物膜结构成分与抗寒性有何关系。 生物膜主要由脂类和蛋白质镶嵌而成,具有一定的流动性,生物膜对低温敏感,其结构成分与抗寒性密切相关。低温下,质膜会发生相变,质膜相变温度随脂肪酸链的加长而增加,随不饱和脂肪酸如油酸、亚油酸、亚麻酸等所占比例的增加而降低,不饱和脂肪酸越多,越耐低温。在缓慢降温时,由于膜脂的固化使得膜结构紧缩,降低了膜对水和溶质的透性;温度突然降低时,由于膜脂的不对称性,膜体紧缩不均而出现断裂,造成膜是破损渗漏,透性加大,胞内溶质外流。生物膜对结冰更为敏感,发生冻害时膜的结构被破坏,与膜结合的酶游离而失去活性。此外,低温也会使膜蛋白质大分子解体为亚基,并在分子间形成二硫键,产生不可逆的凝聚变性,使膜受到伤害。经抗寒锻炼后,由于膜脂中不饱和脂肪酸增多,膜变相的温度降低,膜透性稳定,从而可提高植物的抗寒性。同时,细胞内的NADPH/NADP的比值增高,ATP
糖生物学研究进展
糖生物学研究进展 张文辉 (单位:航天医学工程研究所 学号:w24013 E-mail:pangzizhang503@https://www.360docs.net/doc/4c3467283.html,)摘要:本文主要介绍了糖化学和生物学相结合产生的新学科-糖生物学的概况,主要研究内容、特点及在医学领域中研究动向。 关键词:糖生物学,研究内容,动向。 糖生物学是糖的化学和生物学研究相结合而产生的一门新兴学科,主要研究糖缀合物糖链的结构生物合成和生物学功能,其研究领域包括糖化学、糖链生物合成、糖链在复杂生物系统中的功能和糖链操作技术.糖生物学一经提出,便得到了科学界的广泛认同,并在西方发达国家受到高度重视,在即将到来的后基因组学时代,糖生物学研究更是揭示生命本质所不可缺少的重要方面.已知糖链在细胞内可修饰调控蛋白质、脂类的结构与功能,在细胞外环境参与免疫应答、感染和癌症等过程中的细胞识别但对其作用机制还不完全清楚.近10年来,随着分析技术的进步和分子生物学的发展,糖的研究也取得的了巨大进展,糖生物学研究正成为生命科学研究中又一新的前沿和热点. 糖生物学研究内容: 糖生物学以生物大分子的组成部分糖链为研究对象,研究它作为信息分子在多细胞生物高层次生命活动中的功能,主要包括糖链的结构和功能两个方面的内容。糖链的结构具有惊人的多样性、复杂性和微观不均一性,其一级结构的内容不仅包括各糖基的排列顺序,还包括各糖基的环化形式、各糖基本身异头体的构 型、各糖基间的连接方式以及分支结构的位点和分支糖链的结构。6种单糖形成带分支的六糖有1012异构体。糖链结构的复杂性给糖链的研究带来了很大的困难,同时也使它能携带巨大的生物信息。实际上,糖链的种间特异性、组织特异性以及发育特异性都很强,并且都来源于糖基转移酶不同时间和不同空间的表达。因此,糖基转移酶的研究已成为了当前糖生物学的研究重点。糖复合物中糖链的功能多种多样,如从空间上调节糖复合物的整体结构,保护多肽链不被蛋白酶水解,防止与抗体识别等。近年来的研究表明:糖链作为信息分子涉及多细胞生命的全部空间和时间过程,如精卵识别、组织器官形态形成、老化、癌变等,在血液和淋巴循环中,起着动态的更为灵敏的信号识别和调控作用,涉及到多种严重疾病的发生过程,如炎症和自身免疫病等。关于糖链的生物学作用,有如下一般规则:1)很难预知某一特定的糖链的功能和对生物体的重要性;2)同一寡糖序列在生物体的不同部位和不同的个体发育阶段有不同的功能;3)较为专一的生物作用通常是通过不寻常的序列或常见序列的不寻常表达或修饰来介导的,而这些特殊的糖链也常是毒素和病原体的识别目标。归根结底,糖链的共同特点是介导专一的“识别”和“调控”生物学的过程,因此对糖链的生物学作用也只能逐个地分别研究。当前,糖生物学研究得最多的仍然是糖蛋白。在糖蛋白中,糖链对蛋白质的功能起修饰作用,它通过影响蛋白质的整体构象从而影响由构象决定的所有功能,如蛋白质的正确折叠、细胞内定位、抗原性、细胞-细胞黏附和结合病原体等。在糖脂中人们已经证明了血型的决定物质是糖链,在神经组织及脑中更是存在大量的糖脂,但它们的生理意义至今仍了解得不多。蛋白聚糖主要有维持或抑制细胞生长以及在正常发育和病理条件下结合、贮存及向靶细胞释放生长因子和参与信号转导等作用。细胞表面糖复合物上的糖链是信息功能的承担者,承担着细胞-细胞和细胞-胞外基质的相互作用。[1]
植物生物学试题及答案
植物生物学试题及答案 Document number【SA80SAB-SAA9SYT-SAATC-SA6UT-SA18】
植物学试题一名词解释(12分,2分/个) 1、细胞器; 2、木质部脊; 3、束中形成层; 4、完全叶; 5、花程式; 6、 聚合果 二、判断与改错(17分,对的填“+”,错的填“-”并将错的改正) 1、质体是一类与碳水化合物合成及贮藏相关的细胞器。( ) 2、根毛分布在根尖的伸长区和成熟区。( ) 3、根的初生木质部发育顺序为外始式,而在茎中则为内始式。( ) 4、水生植物叶小而厚,多茸毛,叶的表皮细胞厚,角质层也发达。( ) 5、胡萝卜是变态根,主要食用其次生韧皮部。( ) 6、观察茎的初生结构和次生结构都是从节处取横切面。( ) 7、由3心皮组成的复雌蕊有6条腹缝线。( ) 8、二体雄蕊就是一朵花中只有两个离生的雄蕊。( ) 9、双受精是种子植物的重要特征之一。() 10、桃和梨均为假果。()
三、填空(21分,分/空) 1、植物细胞的基本结构包括和两大部分。后者又可分为、和 三部分。 2、保护组织因其来源及形态结构的不同,可分为和。 3、苎麻等纤维作物所利用的是组织。 4、植物种子是由,和三部分构成,但有些种子却只有和两 部分,前者称种子,后者称种子。 5、禾本科植物的内皮层在发育后期其细胞常面增厚,横切面上,增厚的部分呈形。 6、双子叶植物和裸子植物茎的初生结构均包括、和三部分,玉米等单子叶植 物茎的初生结构包括、和三部分。 7、双子叶植物茎的维管束多为,在初生木质部和初生韧皮部间存在形成层, 故又称维管束。 8、叶的主要生理功能包括和等两个方面。 9、一朵完整的花可分为、、、、和六部分。其中最主 要的部分是和。
糖生物学_植物糖基转移酶研究进展
期末考核 课程:Glycobiology 植物糖基转移酶研究进展 :*** 学号:*** 班级:*** 时间:****
植物糖基转移酶研究进展 摘要:糖基转移酶一类是能够催化糖基从激活的供体转移到特定的受体分子上的一类酶,在生物体中普遍存在并形成了超基因家族。糖基转移酶广泛参与植物生命活动的各种生物学过程。本文综述了近年来的研究报道,综述了糖基转移酶的分类、分离鉴定方法及在生物学功能方面的研究进展,期望为相关研究工作提供参考。 关键词:植物糖基转移酶,分类,分离鉴定,生物学功能 糖基转移酶(Glycosyltransferases,GT,EC 2.4.x.y)是一类催化糖基转移的酶,通过产生糖苷键将供体糖分子或相关基团转移至特异的受体上。糖基转移酶几乎存在于所有的生物体中,其所催化的糖基化反应是最重要的生物学反应之一,直接参与二糖、单糖苷、聚糖苷等的生物合成。糖基供体分子包括双糖、多糖、1-磷酸糖、尿苷二磷酸葡萄糖醛酸,植物中最常见的供体为UDP-Glc。受体可以是糖类、脂类、蛋白质、抗生素和核酸。糖基转移酶催化供体-受体形成α、β两种糖苷键,产物为多糖、糖蛋白、糖脂以及糖苷化合物等。全基因组测序发现真核生物中约1%的基因编码糖基转酶。 1糖基转移酶的分类 目前,对糖基转移酶的分类主要根据Campbell等提出的GT Family 分类系统(数据收录在CAZy数据库中)。糖基转移酶作为高度分歧的多源基因家族,根据蛋白氨基酸序列的一致性、催化特性以及保守序列对其进行分类。因此,一特定的糖基转移酶既可以通过生物化学的方法鉴定其底物,也可以通过生物信息学方法研究其与已知酶基因或酶蛋白氨基酸序列的同源性对其进行分类。目前,依据这种分类方法,糖基转移酶被分为94个家族。根据其的折叠方式可将绝大多数酶分为两个超家族,GT-A超家族和GT-B超家族(图1)。根据催化反应机制、产物的立体化学异构性,在这两个超家族中糖基转移酶又分为反向型和保留型两大类(图2)。 GT-A型折叠的空间结构有两个紧密相连的β/α/β类Rossmann折叠区域。GT-A家族成员需要一个D-X-D基序用来结合二价金金属离子(多为Mn2+),这有助于UDP-糖供体的PPi在酶活性位点上的固定,对于催化反应是不可或缺的。GT-A难以识别UDP-糖供体以外的供体,所以受体的多样性较低。GT-B型折叠的空间含有两个正对的β/α/β类Rossmann折叠区域,连接方式灵活。GT-B成员无需二价金属离子维持活性,这是GT-B与GT-A家族成员的一个显著区别。此外,通过结构分析和PSI-BLAST发现了由跨膜GT组成GT-C超家族,其折叠方式全为反向型,活性位点位于长环部,一般含有8-13个跨膜螺旋。
植物生物学实验教案
植物生物学实验教案授课专业:生物科学、农学主讲:王文龙彭友林李丽 生命科学学院生物基础实验中心二00八年二月十六日
实验1 光学显微镜及体视镜的构造和使用方法 一、实验目的 1、了解光学显微镜及体视镜的一般构造和性能; 2、学会正确地使用光学显微镜及体视镜,熟练地掌握对光,低高倍物镜的使用技术, 以及显微镜的维护; 3、学会临时装片的制作和徒手切片。 二、重点与难点 正确地使用光学显微镜及体视镜,熟练地掌握对光,低高倍物镜的使用技术。三、教学方法与手段 本次课主要采取讲授法和讨论法,在学生实验过程中辅以个别指导进行教学。 四、实验内容 1、光学显微镜的构造、使用方法及维护; 2、临时装片的制作及徒手切片的练习; 3、体视镜的一般结构及使用方法。 五、实验材料 洋葱(Allium cepa)根尖永久装片;洋葱(Allium cepa)鳞片叶;油菜(Brassica campestris)或水稻(Oryza sativa)花粉。 六、实验用品 普通光学显微镜、体视显微镜;镊子、载玻片、盖玻片、培养皿、纱布、吸水纸、擦镜纸、滴瓶、毛笔;碘液、水。 七、实验方法 (一)普通光学显微镜的构造、使用方法及维护 1、显微镜的构造 显微镜的基本结构可以分两部分,即光学部分与机械部分。 (1)光学部分 ①物镜、②目镜、③聚光器、④虹彩光圈、⑤反光镜、⑥镜筒 (2)机械部分 ①镜座、②镜柱、③镜臂、④载物台、⑤物镜转换器、⑥调焦螺旋 2、显微镜的使用方法 (1)正确安置显微镜、(2)对光、(3)低倍物镜的使用、(4)高倍物镜的使用(5)浸油物镜的使用、(6)显微镜的使用练习、(7)用毕复原 3、显微镜的放大倍数 4、光学显微镜的显微测微法 (1)显微测微计
植物生物学试题及答案
植物学试题 一名词解释(12分,2分/个) 1、细胞器; 2、木质部脊; 3、束中形成层; 4、完全叶; 5、花程式; 6、聚合果 二、判断与改错(17分,对的填“+”,错的填“-”并将错的改正) 1、质体是一类与碳水化合物合成及贮藏相关的细胞器。( ) 2、根毛分布在根尖的伸长区和成熟区。( ) 3、根的初生木质部发育顺序为外始式,而在茎中则为内始式。( ) 4、水生植物叶小而厚,多茸毛,叶的表皮细胞厚,角质层也发达。( ) 5、胡萝卜是变态根,主要食用其次生韧皮部。( ) 6、观察茎的初生结构和次生结构都是从节处取横切面。( ) 7、由3心皮组成的复雌蕊有6条腹缝线。( ) 8、二体雄蕊就是一朵花中只有两个离生的雄蕊。( ) 9、双受精是种子植物的重要特征之一。() 10、桃和梨均为假果。() 三、填空(21分,0.5分/空) 1、植物细胞的基本结构包括和两大部分。后者又可分为、和三部分。 2、保护组织因其来源及形态结构的不同,可分为和。 3、苎麻等纤维作物所利用的是组织。 4、植物种子是由,和三部分构成,但有些种子却只有和两部分,前者称种子,后者 称种子。 5、禾本科植物的内皮层在发育后期其细胞常面增厚,横切面上,增厚的部分呈形。 6、双子叶植物和裸子植物茎的初生结构均包括、和三部分,玉米等单子叶植 物茎的初生结构包括、和三部分。 7、双子叶植物茎的维管束多为,在初生木质部和初生韧皮部间存在形成层,故又称维管束。 8、叶的主要生理功能包括和等两个方面。 9、一朵完整的花可分为、、、、和六部分。其中最主要的部分是和。 10、小孢子母细胞进行减数分裂前,花粉囊壁一般由、、和组成。花粉成熟时,花粉囊壁一般 只留下和。 11、被子植物细胞的减数分裂在植物的生活史中发生次。 四、选择题(10分,1分/个) 1、光学显微镜下呈现出的细胞结构称。 A.显微结构;B.亚显微结构;C.超显微结构;D.亚细胞结构 2、裸子植物靠输导水分。 A.导管和管胞;B.管胞;C.筛管;D.筛胞 3、成熟蚕豆种子的种皮上一条黑色眉状物是。 A.种脊;B.种脐;C.种阜;D.种柄 4、原表皮、基本分生组织和原形成层属于。 A.居间分生组织;B.原分生组织;C.初生分生组织;D.伸长区 5、在方形(如蚕豆)或多棱形(如芹菜)的茎中,棱角部分常分布有。 A.厚角组织;B.厚壁组织;C.薄壁组织;D.石细胞 6、栅栏组织属于。 A.薄壁组织;B.分生组织;C.保护组织;D.机械组织 7、以下所列的结构,哪一些都是茎的变态? 。 A.皂荚的分枝刺,葡萄和豌豆的卷须; B.马铃薯、姜、荸荠、芋头; C.莲藕、菊芋、竹鞭、白萝卜、胡萝卜; D.蔷薇和刺槐(洋槐)的刺,豌豆的卷须 8、具的花为整齐花。 A.舌状花冠;B.唇形花冠;C.蝶形花冠;D.十字花冠