RNA聚合酶

在细菌等原核生物中,相同的RNA聚合酶催化三种RNA的合成:信使RNA (mRNA)、核糖体RNA (rRNA)及转运RNA (tRNA)。
细胞RNA聚合酶是相对大的分子。细菌RNA聚合酶是相对大的分子。核心酶有5个亚基(~400 kDa):核心酶有5个亚基(~400 kDa):
α2:这两个α亚基组合成酶及辨认调节因子。每个亚基有两个区,αC末端区及αN末端区,分别与启动子结合及与聚合酶的其他部份结合。每个亚基有两个区,αC末端区及αN末端区,分别与启动子结合及与聚合酶的其他部份结合。
β:有着聚合酶的活动,负责催化RNA的合成。
β':与DNA结合。
ω:还未清楚它的功能。但是它在耻垢分枝杆菌中似乎是提供保护功能予β'亚基。但是它在耻垢分枝杆菌中似乎是提供保护功能予β'亚基。
为着与启动子的特定区域结合,核心酶须有其他亚基,称为σ。为着与启动子的特定区域结合,核心酶须有其他亚基,称为σ。σ因子大大减低RNA聚合酶与非特定的DNA的关系,视乎σ因子本身而增加对某些启动子区域的独特性。σ因子大大减低RNA聚合酶与非特定的DNA的关系,视乎σ因子本身而增加对某些启动子区域的独特性。所以完整的全酶有着6个亚基:α 2 、β、β'、σ及ω(~480 kDa)。所以完整的全酶有着6个亚基:α 2 、β、β'、σ及ω(~480 kDa)。RNA聚合酶的结构就有一个长约55?(即5.5奈米)的沟道及直径为25?(2.5奈米)。RNA聚合酶的结构就有一个长约55?(即5.5奈米)的沟道及直径为25?(2.5奈米)。这个沟道正好适合20?(2奈米)的DNA双股。这个沟道正好适合20?(2奈米)的DNA双股。55?的长度可以接受16核苷酸。55?的长度可以接受16核苷酸。
当不使用时,RNA聚合酶会与弱结合部位结合,等待活性启动子的位点开启并快速转换。当不使用时,RNA聚合酶会与弱结合部位结合,等待活性启动子的位点开启并快速转换。RNA聚合全酶所以在不使用时不是在细胞内自由浮动的。RNA聚合全酶所以在不使用时不是在细胞内自由浮动的。
大肠杆菌的RNA聚合酶组成:
亚基分子量亚基数目功能
α65 000 2 与启动子结合
β15 000 1 含催化部位,起催化作用
β51 000 1 与DNA结合
ω11 000 1
σ70 000 1 识别起始位点
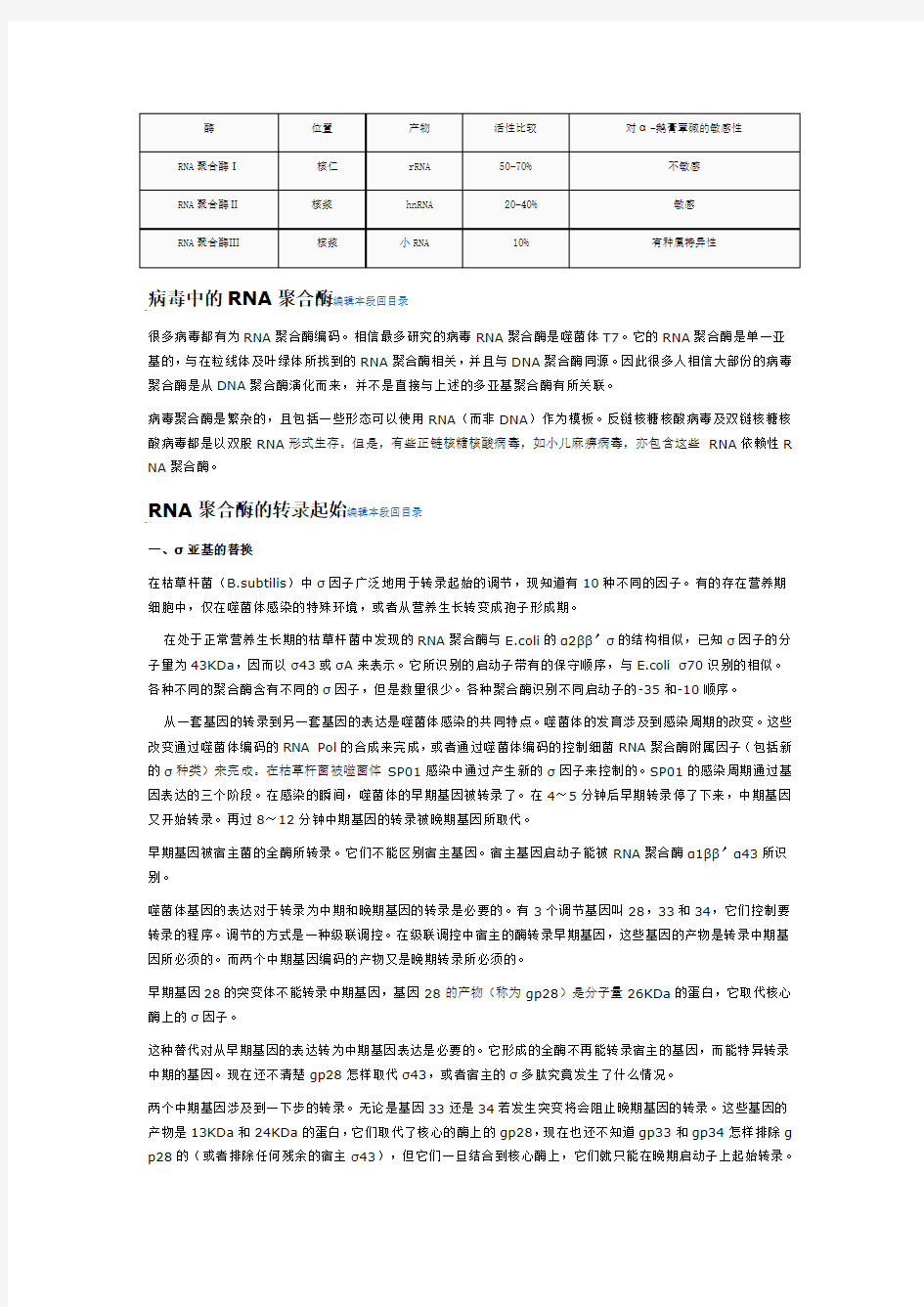
真核生物中三种RNA聚合酶编辑本段回目录
RNA聚合酶I:合成核糖体RNA (rRNA)前体45S,当成熟后会成为28S、18S及5,8S核糖为RNA,是将来核糖体的主要RNA部份。RNA聚合酶I合成核糖体RNA (rRNA)前体45S,当成熟后会成为28S、1 8S及5,8S核糖体RNA,是将来核糖体的主要RNA部份。
RNA聚合酶Ⅱ:合成信使RNA (mRNA)的前体及大部份小核RNA (snRNA)以及微型RNA (microRN A)。RNA聚合酶Ⅱ合成信使RNA (mRNA)的前体及大部份小核RNA (snRNA)以及微型RNA (micr oRNA)。因为它在转录过程中需要多种转录因子才能与启动子結合,所以这是现时最多研究的种类。因为它在转录过程中需要多种转录因子才能与启动子结合,所以这是现时最多研究的种类。
RNA聚合酶Ⅲ:合成转运RNA (tRNAs)、rRNA 5S及其他可以在细胞胞核及原生质找到的細小的RNA。RNA聚合酶Ⅲ合成转运RNA (tRNAs)、rRNA 5S及其他可以在细胞核及原生质找到的细小的RNA。
三种RNA聚合酶的比较:
RNA聚合酶是否有解旋的作用
看2013年浙江省高考题 3.某生物基因表达过程如图所示。下列叙述与该图相符的是 A.在RNA聚合酶作用下DNA双螺旋解开 B.DNA-RNA杂交区域中A应与T配对 C.mRNA翻译只能得到一条肽链 D.该过程发生在真核细胞中 【答案】:A 说明RNA聚合酶有解旋作用 浙科版上有该知识点 这个问题对不同的生物来说不一样,原核生物RNA聚合酶有解旋作用,而对真核生物来说,转录的时候是需要RNA聚合酶和解旋酶一起来发挥解旋作用的。 应该是在转录的过程中,始终没有提到DNA的解旋酶,虽然说酶具有专一性,但是专一性是可以一类的,我问过一些老教师,他们说RNA聚合酶本身有解旋的作用,看到的资料书中,也没有说用DNA解旋酶的。 ... 不同的生物具体情况不同!这是个有争议的话题!也有的书上说,RNA聚合酶属于解旋酶的一种!也有的说RNA聚合酶只能打开起始一段DNA双链,之后需要解旋酶辅助解旋!建议人教版的同学不要再纠结了!个人认为正规考试不会考!仅供参考! 解旋酶一词是在人教版必修一第三章第三节中提出的:“在DNA复制开始时,DNA分子首先利用细胞提供的能力,在解旋酶的作用下,把两条螺旋的DNA链解开,这个过程叫做解旋。”在DNA复制的过程称,解旋需要解旋酶,这是毋庸置疑的。那么转录的时候,DNA的双链解旋也需要解旋酶吗?作者查阅了相关文献资料分别介绍了原核生物的转录起始阶段和真核生物的转录起始阶段。原核细胞内只有一种RNA聚合酶,负责核查3种RNA,RNA酶中的一种因子具有辨认启动子“识别位”并与之结合形成疏松的复合物,然后RNA聚合酶沿模板DNA3'—5'方向移动至Pribnow框(AT含量
RNA聚合酶
在细菌等原核生物中,相同的RNA聚合酶催化三种RNA的合成:信使RNA (mRNA)、核糖体RNA (rRNA)及转运RNA (tRNA)。 细胞RNA聚合酶是相对大的分子。细菌RNA聚合酶是相对大的分子。核心酶有5个亚基(~400 kDa):核心酶有5个亚基(~400 kDa): α2:这两个α亚基组合成酶及辨认调节因子。每个亚基有两个区,αC末端区及αN末端区,分别与启动子结合及与聚合酶的其他部份结合。每个亚基有两个区,αC末端区及αN末端区,分别与启动子结合及与聚合酶的其他部份结合。 β:有着聚合酶的活动,负责催化RNA的合成。 β':与DNA结合。 ω:还未清楚它的功能。但是它在耻垢分枝杆菌中似乎是提供保护功能予β'亚基。但是它在耻垢分枝杆菌中似乎是提供保护功能予β'亚基。 为着与启动子的特定区域结合,核心酶须有其他亚基,称为σ。为着与启动子的特定区域结合,核心酶须有其他亚基,称为σ。σ因子大大减低RNA聚合酶与非特定的DNA的关系,视乎σ因子本身而增加对某些启动子区域的独特性。σ因子大大减低RNA聚合酶与非特定的DNA的关系,视乎σ因子本身而增加对某些启动子区域的独特性。所以完整的全酶有着6个亚基:α 2 、β、β'、σ及ω(~480 kDa)。所以完整的全酶有着6个亚基:α 2 、β、β'、σ及ω(~480 kDa)。RNA聚合酶的结构就有一个长约55?(即5.5奈米)的沟道及直径为25?(2.5奈米)。RNA聚合酶的结构就有一个长约55?(即5.5奈米)的沟道及直径为25?(2.5奈米)。这个沟道正好适合20?(2奈米)的DNA双股。这个沟道正好适合20?(2奈米)的DNA双股。55?的长度可以接受16核苷酸。55?的长度可以接受16核苷酸。 当不使用时,RNA聚合酶会与弱结合部位结合,等待活性启动子的位点开启并快速转换。当不使用时,RNA聚合酶会与弱结合部位结合,等待活性启动子的位点开启并快速转换。RNA聚合全酶所以在不使用时不是在细胞内自由浮动的。RNA聚合全酶所以在不使用时不是在细胞内自由浮动的。 大肠杆菌的RNA聚合酶组成: 亚基分子量亚基数目功能 α65 000 2 与启动子结合 β15 000 1 含催化部位,起催化作用 β51 000 1 与DNA结合 ω11 000 1 σ70 000 1 识别起始位点 真核生物中三种RNA聚合酶编辑本段回目录 RNA聚合酶I:合成核糖体RNA (rRNA)前体45S,当成熟后会成为28S、18S及5,8S核糖为RNA,是将来核糖体的主要RNA部份。RNA聚合酶I合成核糖体RNA (rRNA)前体45S,当成熟后会成为28S、1 8S及5,8S核糖体RNA,是将来核糖体的主要RNA部份。 RNA聚合酶Ⅱ:合成信使RNA (mRNA)的前体及大部份小核RNA (snRNA)以及微型RNA (microRN A)。RNA聚合酶Ⅱ合成信使RNA (mRNA)的前体及大部份小核RNA (snRNA)以及微型RNA (micr oRNA)。因为它在转录过程中需要多种转录因子才能与启动子結合,所以这是现时最多研究的种类。因为它在转录过程中需要多种转录因子才能与启动子结合,所以这是现时最多研究的种类。 RNA聚合酶Ⅲ:合成转运RNA (tRNAs)、rRNA 5S及其他可以在细胞胞核及原生质找到的細小的RNA。RNA聚合酶Ⅲ合成转运RNA (tRNAs)、rRNA 5S及其他可以在细胞核及原生质找到的细小的RNA。 三种RNA聚合酶的比较:
σ因子和RNA聚合酶
σ因子和RNA聚合酶分分合合的关系 在细菌中,如大肠杆菌的RNA聚合酶主要由五个亚基组成,即2αββ,ωσ,其中α亚基与RNA聚合酶的四聚体核心的形成有关,β亚基含有核苷三磷 酸的结合位点,β,亚基含有与DNA模板的结合位点,而σ因子只与RNA转录的起始有关,与链的延伸无关,一旦转录开始,σ因子就被释放,而链的延伸则 由四聚体核心酶催化。 σ因子的特异性决定了它在细菌基因表达转录调控阶段的重要性,即σ因 子和启动子与RNAP的结合具有竞争性,对此,人们提出了σ循环模式,它参与形成延伸复合物,并形成转录延伸的结构基础,其中σ因子在起始和延伸阶段 均发挥作用,但其主要在起始阶段起作用。此循环的主要过程是,①σ因子竞 争性的与核心酶结合形成全酶②全酶与启动子DNA结合形成开放型复合物③σ 因子从延伸阶段的全酶中释放,同时NusA与复合物结合④NusA释放,转录产 物RNA释放,转录终止⑤释放的σ因子又竞争性的与核心酶结合形成σ循环。 这一循环的主要特征是σ因子能快速的重新发挥作用,此循环在生物学在得到 支持,即噬菌体在体外培养时σ因子能与五个或更多的核心酶结合介导转录的 调控。参与此循环的σ因子的数量不定,其数量主要取决于σ合成和降解的速率,以及翻译后的修饰与加工,即其活性与非活性形式之间的转变。此循环的 作用是细胞在面对外界的变化和细胞信号时,此循环能快速的调整转录的形式 以优化细胞的代谢,使用不同启动特异性的σ因子来调控不连续的基因。其中 主要有两大类,第一类是介导管家基因有关的转录,如大肠杆菌中的σ70,第二类主要是其它的因子,如大肠杆菌中的σ32,热激因子只介导合成蛋白质固定和折叠所需的蛋白等。 然而关于σ循环主要有两个版本,第一种是固定性释放,采用色谱或电泳 法比较结合RNAP或延伸复合物的启动子的不同,可以识别结合紧密但易于解离的σ因子,即转录产物RNA-DNA杂交链的长度到达8-9bp之前,σ因子就完全 的解离,此后核心酶参与链的延伸过程,σ因子进入σ循环。第二种是随机释放,发现的机制是改变RNA合成的速度(通过改变转录所需的原料NTP的浓度),但σ因子释放的动力学不变,其主要机理即由于延伸复合物上的σ因子 与核心酶的亲和力降低,核心酶启动转录后σ因子释放就会发生,能降低σ因 子的有效浓度,基于对解离所需时间的运动学的测量,RNA合成的速率不会影 响σ因子解离的运动学时间,新生的RNA对σ因子的解离也无影响,解离是相 对随机,此种机制中可能发生第一种版本,即固定性释放。此外还有一些其他 大的版本,如σ因子不解离或部分解离全酶纯化技术,能用来解释大多数的 转录在指数期,而σ70介导的转录大部分在稳定期,同样这也引发了一些思考,比如,因子的保留存在于体内还是在细胞溶解和RNAP纯化过程中形成的。 因子释放和保留的结构基础,与管家基因有关的σ因子被分成四个保守的序列 区,它们又被分为4个独立的结构域即σ 1.1 σ 2 σ 3 和σ 4 ,相邻的结构域灵活连 接,它们与RNAP至少有五个不连续的结合区,如,σ 1.1 位于或靠近于下游的 DNA通道,σ 2 与β,形成卷曲螺旋的结构,这些相互作用既能稳定也能破坏百个以上氨基酸链的相互作用。因子与RNAP不同部分之间的同时但又各自独立的结合,增加了σ因子与酶的亲和力,但会引起结构的改变,单个σ因子不同结
真核生物三种RNA聚合酶的特点
1.简述真核生物三种RNA聚合酶的特点? 下边是详细的 RNA聚合酶Ⅰ的转录产物是45SrRNA,经剪接修饰后生成除5SrRNA 外的各种rRNA。rRNA与蛋白质组成的核糖体是蛋白质合成的场所。RNA聚合酶Ⅱ在核内转录生成hnRNA,经剪接加工后生成的mRNA被运送到胞质中作为蛋白质合成的模板。 RNA聚合酶Ⅲ的转录产物是tRNA,5SrRNA,snRNA,其中snRNA参与RNA的剪接。 2.简述乳糖操纵子的调控原理? 答:答:(1)乳糖操纵子的组成:大肠杆菌乳糖操纵子含Z、Y、A三个结构基因,分别编码半乳糖苷酶、透酶和半乳糖苷乙酰转移酶,此外还有一个操纵序列O,一个启动子P和一个调节基因I. (2)阻遏蛋白的负性调节:没有乳糖存在时,I基因编码的阻遏蛋白结合于操纵序列O处,乳糖操纵子处于阻遏状态,不能合成分解乳糖的三种酶;有乳糖存在时,乳糖作为诱导物诱导阻遏蛋白变构,不能结合于操纵序列,乳糖操纵子被诱导开放合成分解乳糖的三种酶。所以,乳糖操纵子的这种调控机制为可诱导的负调控。(3)CAP的正调节:在启动子上游有CAP结合位点,当大肠杆菌从以葡糖糖为碳源的环境转变为以乳糖为碳源的环境时,cAMP浓度升高,与CAP结合,使CAP发生变构,CAP结合于乳糖操纵子启动序列附近的CAP结合位点,激活RNA聚合酶活性,促进结构基因转录,调节蛋白结合于操纵子后促进结构基因的转录,对乳糖操纵子实行正调控,加速合成分解乳糖的三种酶。 (4)协调调节:乳糖操纵子中的I基因编码的阻遏蛋白的负调控与CAP的正调控两种机制,互相协调、互相制约。 3.简述DNA聚合酶和DNA连接酶在DNA复制中的作用? 答:DNA聚合酶(DNA polymerase)是细胞复制DNA的重要作用酶。 DNA聚合酶 , 以DNA为复制模板,从将DNA由5'端点开始复制到3'端的酶。 DNA聚合酶的主要活性是催化DNA的合成(在具备模板、引物、dNTP等的情况下)及其相辅的活性。 真核细胞有5种DNA聚合酶,分别为DNA聚合酶α(定位于胞核,参与复制引发具5-3外切酶活性),β(定位于核内,参与修复,具5-3外切酶活性),γ(定位于线粒体,参与线粒体复制具5-3和3-5外切活性),δ(定位核,参与复制,具有3-5和5-3外切活性),ε(定位于核,参与损伤修复,具有3-5和5-3外切活性)。 原核细胞有3种DNA聚合酶,都与DNA链的延长有关。DNA聚合酶I是单链多肽,可催化单链或双链DNA 的延长;DNA聚合酶II 则与低分子脱氧核苷酸链的延长有关;DNA聚合酶III在细胞中存在的数目不多,是促进DNA链延长的主要酶。 DNA连接酶是1967年在三个实验室同时发现的,最初是在大肠杆菌细胞中发现的。它是一种封闭DNA链上缺口酶,借助ATP或NAD水解提供的能量催化DNA链的5'-PO4与另一DNA链的3'-OH 生成磷酸二酯键。但这两条链必须是与同一条互补链配对结合的(T4DNA连接酶除外),而且必须是两条紧邻DNA链才能被DNA连接酶催化成磷酸二酯键。DNA连接酶在大肠杆菌细胞中约有300个分子,和DNA聚合酶Ⅰ的分子数相近,这也是比较合理的现象。因为DNA连接酶的主要功能就是在DNA聚合酶Ⅰ催化聚合,填满双链DNA 上的单链间隙后封闭DNA双链上的缺口。这在DNA复制、修复和重组中起着重要的作用,连接酶有缺陷的突变株不能进行DNA复制、修复和重组。 4.简述真核生物基因时空表达的意义? 答:所有生物的基因表达都具有严格的规律性,即表现为时间特异性和空间特异性。 基因表达的时间特异性是指某一特定基因的表达按一定的时间顺序发生。如编码甲胎蛋白的基因在胎儿肝细胞中活跃表达,因此合成大量的甲胎蛋白。在成年后该基因的表达水平很低,几乎检测不到ATP。但是,当肝细胞发生转化形成肝癌细胞时,编码ATP的基因又重新被激活,大量的AFP被合成。 空间特异性是指多细胞生物个体在某一特定生长发育阶段,同一基因在不同的组织器官表达不同。在多细胞生物个体某一发育、生长阶段,同一基因产物在不同的组织器官表达水平是不一样的。在个体生长、发育过程中,一种基因产物在个体的不同组织或器官表达,即在个体的不同空间出现,这就是基因表达的空间特异性。 5.已知一个编码蛋白质的cDNA序列,如何才能得到相应的蛋白质? 1、从基因文库或cDNA文库中分离目的基因或通过pcr扩增得到目的基因; 2、体外构建重组DNA分子,选择合适的克隆载体如PBV220,用限制型内切酶分别加工外源基因与载体DNA,再用T4DNA连接酶连接外源基因与载体DNA,构建出重组质粒DNA分子; 3、重组质粒DNA导入宿主细胞:制备感受态大肠杆菌宿主细胞DH5α,将重组题分子导入宿主细胞,涂布在含有Amp的LB固体平板上,37℃培养; 4、挑选单菌落,提取质粒,用限制性内切酶酶切鉴定,如果酶切产物的大小为2.7kb左右,则通过测序确认序列的正确性; 5、表达:将所得阳性克隆在30℃培养适当时间,使得菌密度较高以后,升温42℃诱导基因的表达,2~3小时以后,离心收集菌体,用SDS-PAGE检测目的基因的表达; 6、纯化:破细胞后用适当的方法纯化出目的蛋白。 6.结合你的课题,举例说明你的硕士阶段可能会用到的3种分子生物学技术(如果研究领域不涉及分子生物学技术,可简单谈谈自己研究领域的前景) 答:(1)Western Blot,所谓免疫印迹是一种将凝胶电泳与固相免疫结合起来的分子生物学技术。就是把正常的细胞打碎,分离出